Download presentation
1
第八章 DNA的复制和修复 第一节 DNA的复制 第二节 DNA的损伤及修复

2
信息传递的中心法则 复制 DNA 转录 调控 逆转录 调控 蛋白质 RNA 翻译 复制

3
第一节 DNA的复制 一、 DNA复制的基本规律 二、 DNA复制中所需的酶和辅因子 三、原核生物的DNA复制过程

4
T4噬菌体DNA

5
一、DNA复制的基本规律

7
(一)DNA的复制方式——半保留复制 1、半保留复制概念:在DNA复制过程中,亲代的一个DNA双螺旋分子通过复制形成了两个与原先的碱基序列完全相同的子代DNA分子,每个子代分子中有一条链来自亲代DNA,另一条链是新合成的。这样的复制方式,称为半保留复制。
DNA的复制方式——半保留复制 1、半保留复制概念:在DNA复制过程中,亲代的一个DNA双螺旋分子通过复制形成了两个与原先的碱基序列完全相同的子代DNA分子,每个子代分子中有一条链来自亲代DNA,另一条链是新合成的。这样的复制方式,称为半保留复制。")
8
2、DNA半保留复制的证据 15N-DNA的密度大于14N-DNA的密度

9
(二)DNA复制的模式 复制子(replicon):基因组中能独立进行复制的单位。
起点(origin of replication ,ori):以一条链为模板,起始合成DNA的一段序列,富含A.T。 每个复制子都含有一个复制起点。 原核生物是单复制子,真核生物是多复制子 终点(terminus,ter):与复制终止有关位点。 其中已发现Ecoli的复制终点含有23bp的保守序列,由tus蛋白与此位点结合参与复制的终止。 真核生物中似乎没有复制终止点
:以一条链为模板,起始合成DNA的一段序列,富含A.T。 每个复制子都含有一个复制起点。 原核生物是单复制子,真核生物是多复制子. 终点(terminus,ter):与复制终止有关位点。 其中已发现Ecoli的复制终点含有23bp的保守序列,由tus蛋白与此位点结合参与复制的终止。 真核生物中似乎没有复制终止点.")
10
复制起点 83 33 复制终点 E.Coli基因结构和复制起止点

11
复制的方向:单向或双向 双向 单向

13
原核生物的θ复制:双向复制 复制叉

14
复制叉 起点 延伸 领头链 随后链 3’ 5’ θ形复制

16
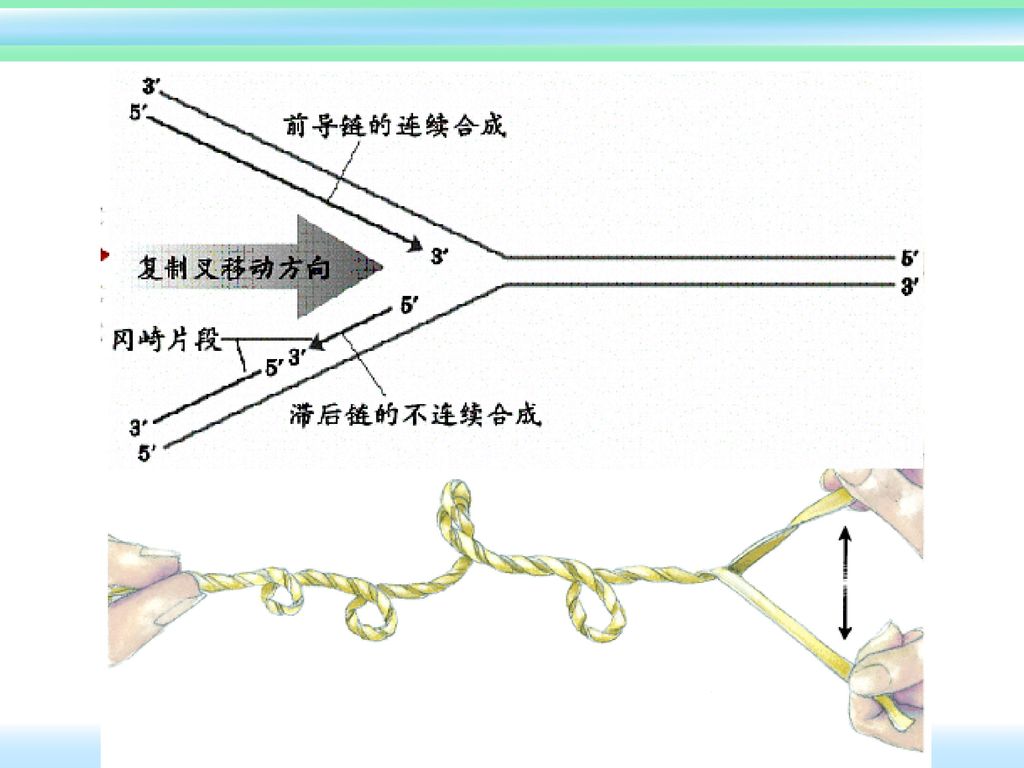
三、 半不连续复制 半不连续复制—在DNA复制时,领头链是连续合成的,而随从(滞后)链的合成是不连续的,这种复制方式称为半不连续复制。
不管是DNA还是RNA,核酸链的合成方向都是5’→3’ 前导链(领头链):一条链以走向3’→5’的亲代链为模板,能连续合成,合成方向与复制叉移动方向同向,称为~。 滞后链(随从链):另一条链以走向5’→3’的亲代链为模板时,只能按5’→3’的方向合成许多不连续的小片段(称为冈崎片段),最后再连成一条完整的子代DNA链。合成方向与复制叉移动方向相反,称为~.
:一条链以走向3’→5’的亲代链为模板,能连续合成,合成方向与复制叉移动方向同向,称为~。 滞后链(随从链):另一条链以走向5’→3’的亲代链为模板时,只能按5’→3’的方向合成许多不连续的小片段(称为冈崎片段),最后再连成一条完整的子代DNA链。合成方向与复制叉移动方向相反,称为~.")
17
3’ 5’ Rep蛋白 解链酶 SSB 前导链 滞后链

18
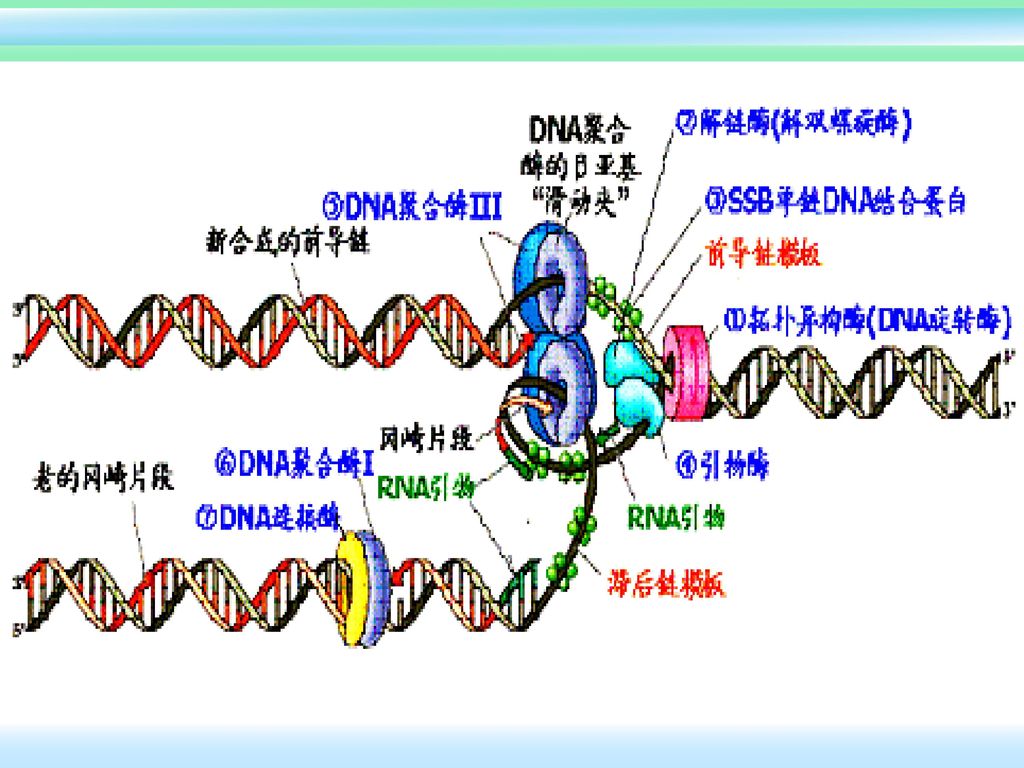
四、DNA复制中所需的酶和辅因子 模板DNA 前导链模板 解旋酶 引物 后滞链模板 DNpolymerase 连接酶 冈崎片段 新合成的
已连接的 单链结合蛋白 新合成的 前导链 DNA聚合酶

19
(一)、DNA聚合酶 1、在5’端有RNA(或DNA)存在的前提下,延长DNA子链 作用特点:不能从头合成子链,只能延长子链
(1)以四种脱氧核苷酸三磷酸为底物; (2)反应需要有DNA模板的指导 (3)反应需要有引物3-OH存在; (4)DNA链的合成方向为5 3 ; (5)产物DNA的性质与模板相同 (6)DNA的体外聚合必须加入少量的DNA才能进行。
以四种脱氧核苷酸三磷酸为底物; (2)反应需要有DNA模板的指导. (3)反应需要有引物3-OH存在; (4)DNA链的合成方向为5 3 ; (5)产物DNA的性质与模板相同. (6)DNA的体外聚合必须加入少量的DNA才能进行。")
20
2、原核生物中的DNA聚合酶 E.COLi的DNA聚合酶 功 能 Ⅰ Ⅱ Ⅲ 5’→3’聚合酶作用 + + +
功 能 Ⅰ Ⅱ Ⅲ 5’→3’聚合酶作用 5’→3’外切酶作用 - - 3’→5’外切酶作用 聚合核苷酸数 /min 分子数/细胞 *DNApolⅢ主要负责DNA链延伸。

21
DNA聚合酶的功能: 5’→3’聚合酶的活性催化dNTP加到DNA链的3’-OH末端(方向5’ →3’)
5’ →3’外切酶活性,作用于双链DNA的碱基配对部分,从5’端切下单核苷酸或寡核苷酸。在切除RNA引物中发挥作用。 3’ → 5’外切酶活性,作用于单链DNA3’末端核苷酸,能及时切除错配核苷酸,起校对功能,负责DNA的损伤修复

22
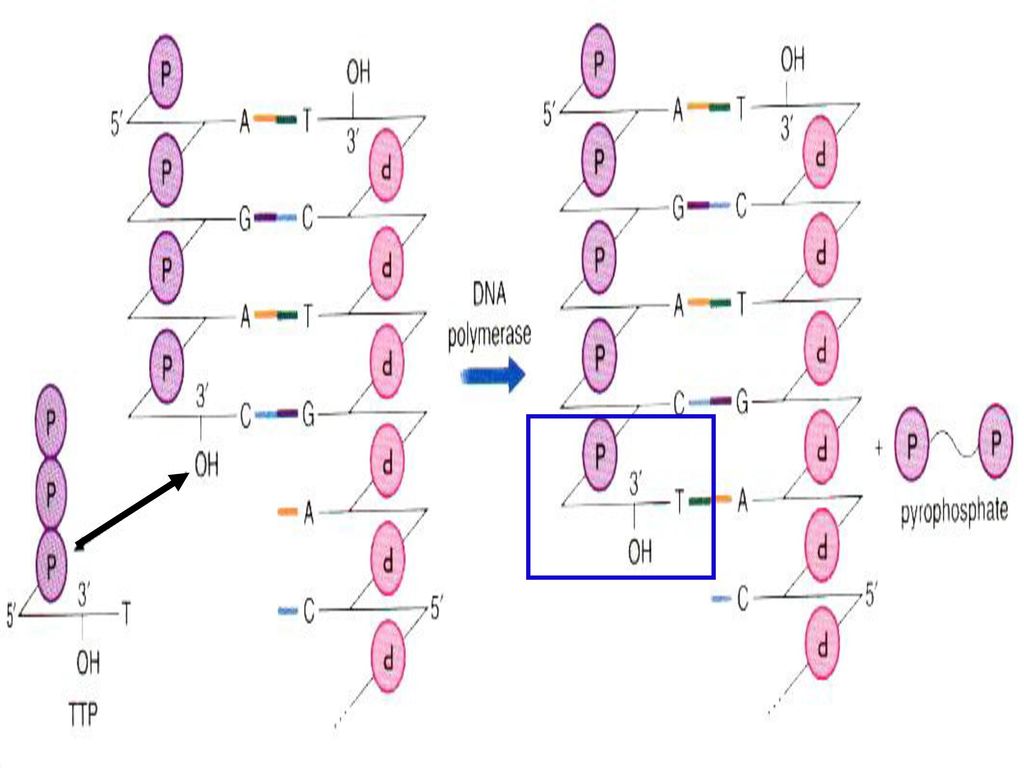
DNA聚合酶的5’→3’聚合酶的活性 P O H 5’ 5 3 + ¡ä ’ DNA 聚合酶 - α β γ PPi

24
3、真核生物的DNA聚合酶(DNA-pol):
无

25
(二)引物酶 引物酶:此酶以DNA为模板合成一小段RNA。实质是以DNA为模板的RNA聚合酶。
⑴从模板复制最初几个核酸时,碱基堆集力和氢键都较弱,易发生错配 ⑵新复制的最初几个核苷酸,没有与模板形成稳定双链,DNA聚合酶的5’→3’校对功能难发挥作用

27
(三)DNA连接酶(ligase,1967年发现)
作用: 催化双链DNA中一条链上缺口的共价连接,形成3’,5’-磷酸二酯键 条件: ①两片段相邻;②两片段需与同一互补链结合; ③反应耗能:细菌要求NAD+,动物和噬菌体要求ATP提供能量。

29
(四)DNA解螺旋酶(解链酶) 作用:通过水解ATP来断裂双链DNA互补碱基对之间的氢键,使DNA双螺旋分离形成复制叉
每解1个bp需耗2个ATP

30
(五)DNA拓扑异构酶 催化DNA的拓扑连环数发生变化的酶。 作用:解开DNA超螺旋,改变连环数 螺旋 超螺旋
DNA拓扑异构酶 催化DNA的拓扑连环数发生变化的酶。 作用:解开DNA超螺旋,改变连环数 螺旋 超螺旋")
31
DNA拓扑异构酶改变连环数

32
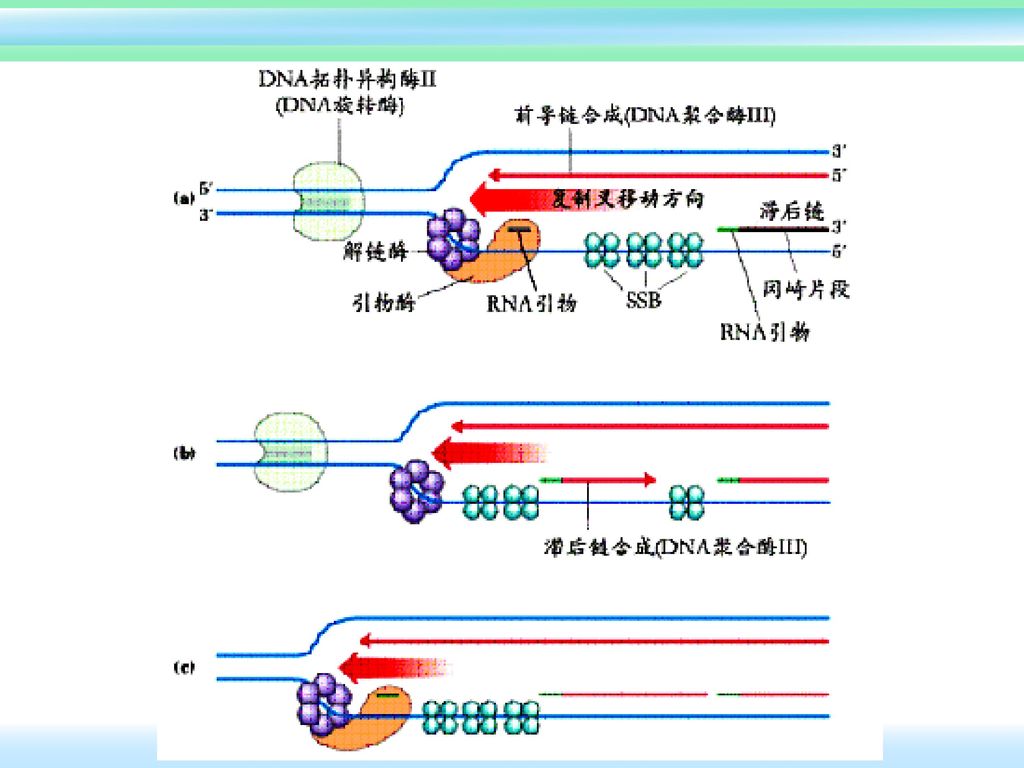
拓扑异构酶Ⅰ 每次切断DNA双链中的一股和再连接 反应不需要能量。 主要集中在活性转录区,同转录有关。 拓扑异构酶Ⅱ 可同时切断DNA双链,解旋或再螺旋后再连接.兼内切酶和连接酶活性。 需要能量(ATP或NAD+) 有ATP时可引入2个负超螺旋。引入负超螺旋可消除复制叉前进带来的张力,促进解链,同复制有关。

33
DNA拓扑异构酶Ⅱ的作用机制

34
(Single-strand binding protein, SSB):
(六)单链结合蛋白 (Single-strand binding protein, SSB): 结合在已解开的DNA单链上 作用: ①保护单链DNA免遭核酸酶的降解,使单链DNA保持伸展态,以便作为模板。 ②降低DNA的Tm,促进DNA解链,阻止复性。
单链结合蛋白. (Single-strand binding protein, SSB): 结合在已解开的DNA单链上. 作用: ①保护单链DNA免遭核酸酶的降解,使单链DNA保持伸展态,以便作为模板。 ②降低DNA的Tm,促进DNA解链,阻止复性。")
35
DNA复制酶 系一览

37
四.原核生物的DNA复制过程 1、复制的起始 原核生物从一个固定的起始点开始,同时向两个方向进行的双向复制——θ复制,复制起点都是富含A、T的区段。 大肠杆菌复制起点:oriC 终止点:ter 复制叉 终止区ter

38
E.coli复制起点oriC跨度为245bp,包含有三组串联重复序列和两对反向重复序列

39
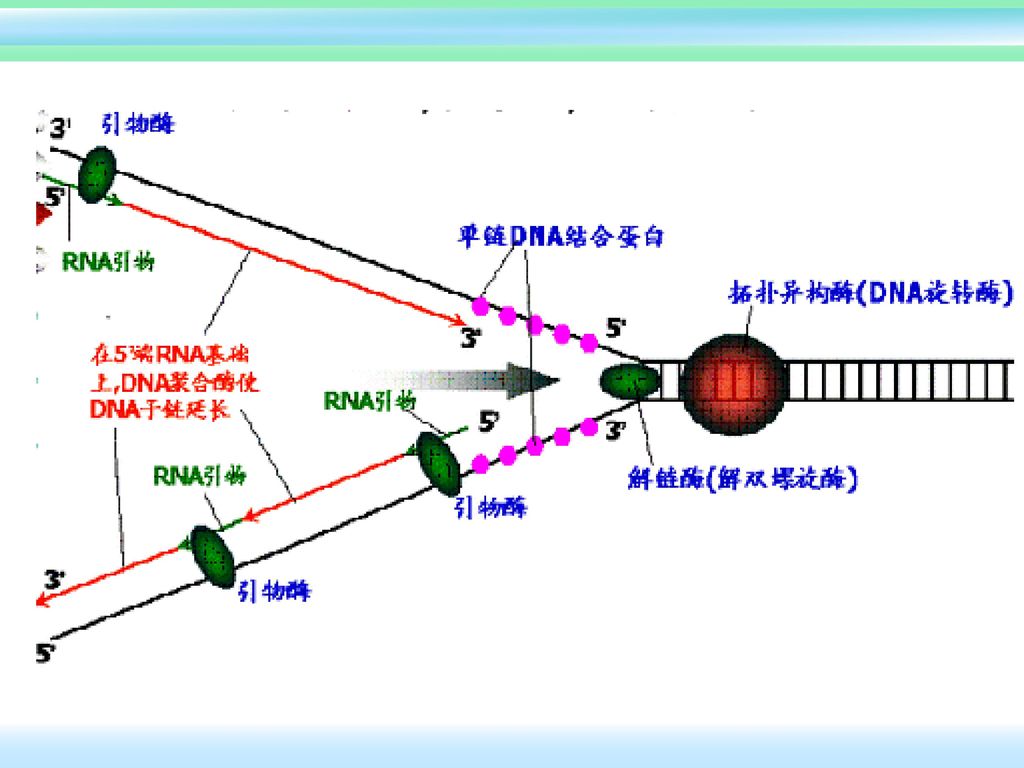
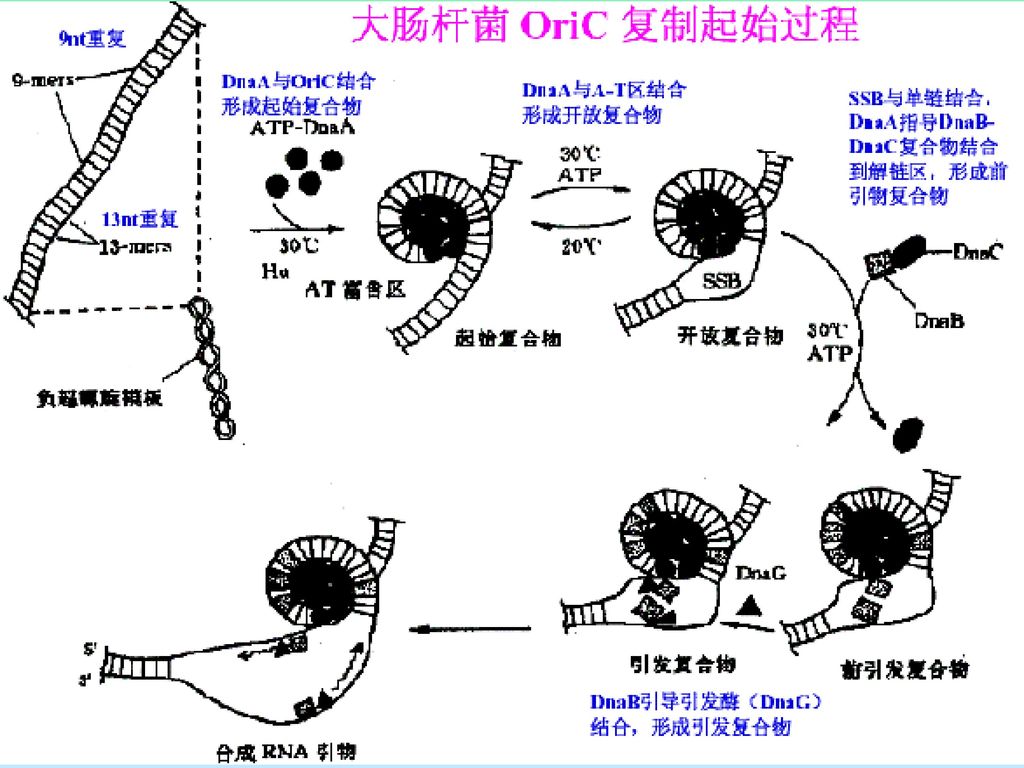
起始过程: 一些特殊的蛋白可以识别并结合到复制起点,DNA环绕此复合物 使DNA双螺旋局部解链,在起始位点处形成复制眼、复制叉。 由引发合成酶合成引物。滞后链需不断合成冈崎片段的RNA引物 复制起点的识别 DNA解链 RNA引物的合成

40
大肠杆菌复制原点起始复制所需蛋白质: DNaA 识别起点序列,在起点处打开双螺旋 DNaB 使DNA解旋 DNaC DNaB结合在原点所需 Hu 刺激起始 引物酶(DNaG) 合成RNA引物 SSB 结合单链DNA RNA聚合酶 促进DNaA活性 旋转酶(TOPOⅡ) 松驰DNA扭曲应力
 合成RNA引物. SSB 结合单链DNA. RNA聚合酶 促进DNaA活性. 旋转酶(TOPOⅡ) 松驰DNA扭曲应力.")
42
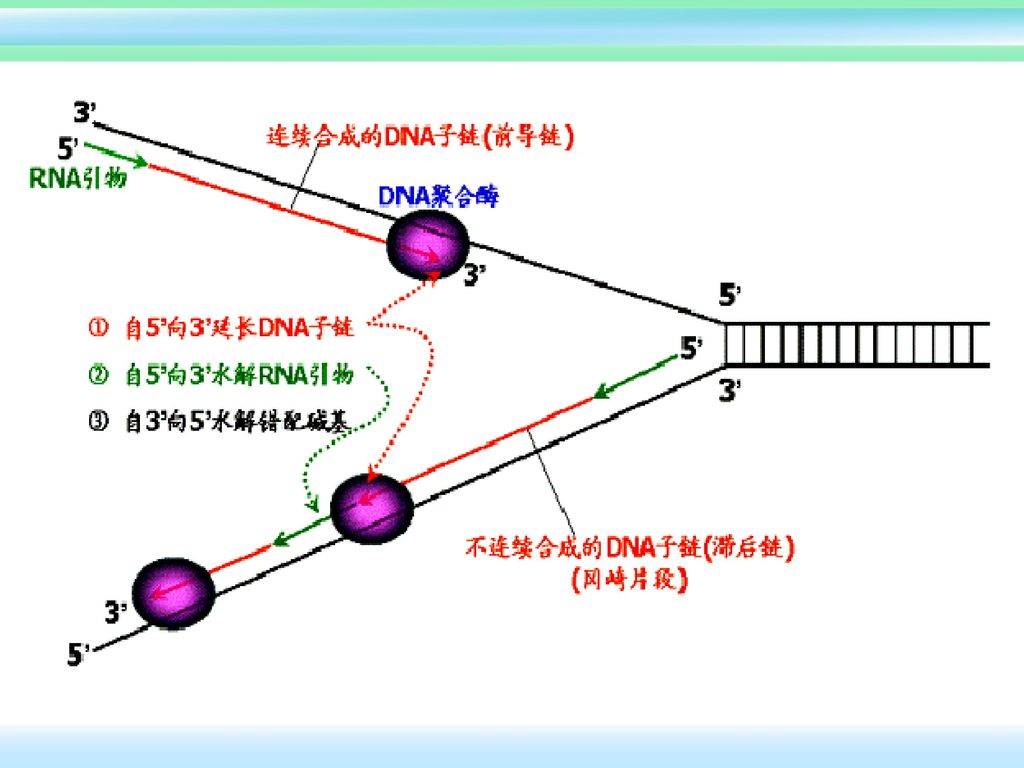
2、DNA链的合成与延伸 前导链和滞后链的延伸同时进行。
子链延伸:引物合成后,在DNA聚合酶Ⅲ作用下,自RNA引物3’-OH末端依次添加与模板链对应互补的新的脱氧核苷酸。 半不连续合成: (1)前导链的延伸:连续合成,延伸方向与解链方向相同 (2)滞后链的合成:是分段进行的,延伸方向与解链方向相反。 不断合成冈崎片段的RNA引物,由DNA聚合酶Ⅲ按5’→3’的方向合成许多冈崎片段;当新形成的冈崎片段延长至一定长度,其3’-OH端遇到上一个冈崎片段时即停止合成。
前导链的延伸:连续合成,延伸方向与解链方向相同. (2)滞后链的合成:是分段进行的,延伸方向与解链方向相反。 不断合成冈崎片段的RNA引物,由DNA聚合酶Ⅲ按5’→3’的方向合成许多冈崎片段;当新形成的冈崎片段延长至一定长度,其3’-OH端遇到上一个冈崎片段时即停止合成。")
44
3、复制终止 复制叉移动到终止区即停止复制:
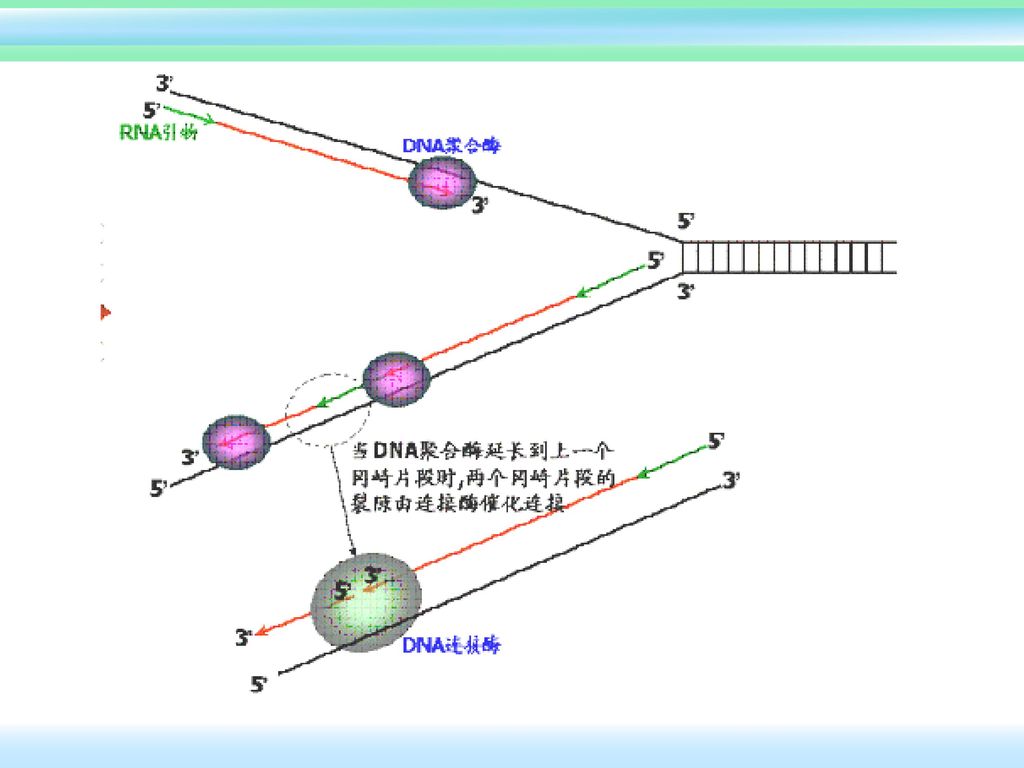
切除引物,补齐缺口:由DNA聚合酶Ⅰ(5’→3’外切活性)催化,切去RNA引物;按碱基互补原则,沿5’→3’方向,留下的空隙由DNA聚合酶Ⅰ(5’→3’合成活性)催化合成一段DNA补齐缺口。 连接封口:由DNA连接酶催化,将补齐缺口的3’-OH基与下一个冈崎片段的5’-P以磷酸二酯键连接起来。 校正并修复DNA:由DNA聚合酶校正并切除错配,再按5’→3’方向加上正确核苷酸。以修复方式填补终止区50-100bp的空缺。
催化,切去RNA引物;按碱基互补原则,沿5’→3’方向,留下的空隙由DNA聚合酶Ⅰ(5’→3’合成活性)催化合成一段DNA补齐缺口。 连接封口:由DNA连接酶催化,将补齐缺口的3’-OH基与下一个冈崎片段的5’-P以磷酸二酯键连接起来。 校正并修复DNA:由DNA聚合酶校正并切除错配,再按5’→3’方向加上正确核苷酸。以修复方式填补终止区50-100bp的空缺。")
45
5’ 3’ 3’ 5’ 滞后链的合成 Pol Ⅲ 5’ 3’ DNA Pol I 5’ 3’ DNA 连接酶 5’ 3’ 5’ 3’

47
DNA复制时酶的作用次序: ⑴ DNA解螺旋酶解开双链DNA。 ⑵ SSB结合于DNA单链。
复制叉前进时带来的扭曲张力。 ⑷ DNA引物酶(在引发体中)合成RNA引物。 ⑸ DNA polⅢ在两条新生链上合成DNA。 ⑹ DNA polⅠ切除RNA引物,并补上DNA。 ⑺ DNA ligase连接两个冈崎片段。
合成RNA引物。 ⑸ DNA polⅢ在两条新生链上合成DNA。 ⑹ DNA polⅠ切除RNA引物,并补上DNA。 ⑺ DNA ligase连接两个冈崎片段。")
48
五、真核细胞DNA的复制 1、真核生物中DNA的复制特点 真核生物染色体是线形的 有多复制子(多起点双向复制)。
真核生物染色体在全部复制完之前起点不再重新开始复制;而在快速生长的原核中,起点可以连续发动复制。 真核生物在DNA复制的同时另外还要组装成核小体,组蛋白则是全保留的。 真核生物线性染色体两端有端粒结构,防止染色体间的末端连接。由端粒酶负责新合成链5RNA引物切除后的填补,亦保持端粒的一定长度。

49
2、真核生物的端粒和端粒酶 DNA复制导致末端缩短 DNA聚合酶切除单链DNA 5’ 3’ 引物 引物 DNA聚合酶切除引物 5’ 3’

50
端粒(telomere) 是真核生物染色体线性DNA分子末端特殊的结构 ,富含GC的重复序列 人:(TTAGGG)n
端粒酶(telomerase) 催化端粒复制的一种核糖核蛋白,携带RNA模板(与端粒互补)的逆转录酶;既有模板,又有逆转录酶的作用 端粒结构 3 5 功能: ⑴保证线性DNA的完整复制 ⑵保护染色体末端 ⑶决定细胞寿命,胚系细胞含端粒酶,体细胞不表达端粒酶。肿瘤细胞端粒酶活性恢复
 催化端粒复制的一种核糖核蛋白,携带RNA模板(与端粒互补)的逆转录酶;既有模板,又有逆转录酶的作用. 端粒结构. 3 5 功能: ⑴保证线性DNA的完整复制. ⑵保护染色体末端. ⑶决定细胞寿命,胚系细胞含端粒酶,体细胞不表达端粒酶。肿瘤细胞端粒酶活性恢复.")
51
端粒复制: 1)借本身RNA与末端ssDNA互补; 2)以酶上RNA为模板延伸ssDNA; 3)延长的ssDNA又可反折为引物,合成双链。
借本身RNA与末端ssDNA互补; 2)以酶上RNA为模板延伸ssDNA; 3)延长的ssDNA又可反折为引物,合成双链。")
52
第二节 DNA的损伤及修复 一、 DNA的损伤: 破坏DNA的双螺旋结构,造成DNA结构和功能的破坏。 UV造成的基因改变 嘧啶二聚体的形成

53
互变异构:A=NH时可形成A=C,G-OH时可形成GT三键配对 碱基脱氨:C-U,A-I,G-X
二、 DNA的损伤原因: 自发性损伤 复制时的碱基错配 互变异构:A=NH时可形成A=C,G-OH时可形成GT三键配对 碱基脱氨:C-U,A-I,G-X 碱基丢失:大肠杆菌每代丢失一个嘌呤,哺乳动物可达一万个。嘧啶丢失几率只有嘌呤的1∕20。

54
环境作用引起的损伤: 诱变因素及突变类型

55
三、 损伤的修复 直接修复 切除修复 重组修复 诱导修复
细胞内具有一系列起修复作用的酶系统,可以除去DNA上的损伤,恢复DNA的双螺旋结构。目前已知有4种酶修复系统: 直接修复 切除修复 重组修复 诱导修复

56
1、直接修复 (1)光复活修复: 可见光激活了光复活酶,它能分开由于UV照射形成的TT(CC CT)嘧啶二聚体,而恢复原来状态。
光复活修复: 可见光激活了光复活酶,它能分开由于UV照射形成的TT(CC CT)嘧啶二聚体,而恢复原来状态。")
57
紫外线损伤的光复活过程:A 形成嘧啶二聚体,B. 光复合酶结合于损伤部位, C 酶被可见光激活,D. 修复后释放酶
光修复酶可使二聚体解聚为单体状态,DNA完全恢复正常。 光修复酶的激活需 μm波长的光。 光修复酶广泛存在,对单细胞生物比较重要,但人体只存在于淋巴细胞和皮肤成纤维细胞,且是次要修复方式。

58
(2)O6-甲基鸟嘌呤的修复 甲基化的鸟嘌呤在甲基转移酶的作用下,将甲基转移到酶自身的半胱氨酸残基上
O6-甲基鸟嘌呤的修复 甲基化的鸟嘌呤在甲基转移酶的作用下,将甲基转移到酶自身的半胱氨酸残基上")
59
2、切除修复: 在一系列酶作用下, 将DNA受损部分切 掉,并以完整链为模 板,合成出被切部分, 从而使DNA恢复正

60
I、结构缺陷的核苷酸切除修复: (1)核酸内切酶识别DNA损伤部位,在其附近将其切开。 (2)核酸外切酶切除损伤的DNA。 (3)DNA聚合酶修复。 (4)DNA连接酶连接。
核酸内切酶识别DNA损伤部位,在其附近将其切开。 (2)核酸外切酶切除损伤的DNA。 (3)DNA聚合酶修复。 (4)DNA连接酶连接。")
61
II、碱基切除修复 DNA糖苷酶可识别不正常的碱基,水解下不正常的碱基,形成AP(无嘌呤无嘧啶)位点 对于AP位点的损伤的修复方法:
核酸外切酶切除, DNA聚合酶修复, DNA连接酶连接。

62
3、重组修复 当DNA发动复制时尚未修复的损伤部位,可以先复制,再重组修复。(发生在复制后)
")
63
4、诱导修复 DNA严重损伤能引起一系列复杂的诱导效应,称为应急反应(SOS response)。
倾向差错的修复:SOS反应还能诱导产生缺乏校对功能的DNA聚合酶,它能在DNA损伤部位进行复制而避免了死亡,可是却带来了高的突变率,这属于倾向差错的修复。是在DNA损伤面太大,复制难以继续的情况下发生。特异性很低。

64
本章要求 1、掌握生物学中心法则、半保留复制,不连续复制、逆转录酶的概念。 2、掌握DNA聚合酶催化的反应,复制保真性依赖的机理。











 考点二: 细胞膜与细胞壁(实验 “ 验证活细胞 吸收物质的选择性 ” :目的要求、材料用具、方 法步骤、实验现象和结果、讨论) 考点三:细胞质(实验 “ 观察叶绿体的形态和分 布.>")

![学案6 基础实验 [题型剖析] 教材中基础实验的考查是近几年试题命制的一个趋势,本类试题主要考查考生对相关实验的目的、原理、方法、操作步骤和相关技能的掌握情况。主要分为显微观察类、物质鉴定类、探究设计类、调查模拟类。 [突破策略] 首先要将教材中的实验分类归纳;然后对教材中每个实验的目的、原理、材料、步骤及注意事项等进行比较记忆,要认真领悟每个实验的设计意图,并从其中提炼总结实验方法和技术。](/60/11174577/big_thumb.jpg "学案6 基础实验 [题型剖析] 教材中基础实验的考查是近几年试题命制的一个趋势,本类试题主要考查考生对相关实验的目的、原理、方法、操作步骤和相关技能的掌握情况。主要分为显微观察类、物质鉴定类、探究设计类、调查模拟类。 [突破策略] 首先要将教材中的实验分类归纳;然后对教材中每个实验的目的、原理、材料、步骤及注意事项等进行比较记忆,要认真领悟每个实验的设计意图,并从其中提炼总结实验方法和技术。>")

 南京市第三高级中学 周敏.>")






>")
