Download presentation
Presentation is loading. Please wait.

1
Chapter 16 Oncogene, Anti-oncogene and Growth Factor
第十六章 癌基因、抑癌基因与生长因子 Chapter Oncogene, Anti-oncogene and Growth Factor

2
引言 据估计,大约有三分之一的人在生命的某一阶段要遭遇癌症。
迄今发现的癌症有200多种,我国最为常见和危害性最严重的肿瘤为肺癌、食管癌、胃癌、大肠癌、肝癌、乳腺癌、鼻咽癌、宫颈癌和淋巴瘤、白血病。 根据卫生部肿瘤防治办公室提供的2006年我国肿瘤发病率和十大恶性肿瘤发病率排序显示,肺癌、乳腺癌分别位居男、女性恶性肿瘤发病首位,男女恶性肿瘤死亡率最高的均为肺癌。

3
根据肿瘤的生物学特性及其对机体的危害性的不同,肿瘤可分为良性肿瘤和恶性肿瘤两大类。
肿瘤(tumor)是指细胞在致癌因素作用下,基因发生了改变,失去对其生长的正常调控,导致单克隆性异常增生而形成的新生物。这种新生物常形成局部肿块,因而得名。 根据肿瘤的生物学特性及其对机体的危害性的不同,肿瘤可分为良性肿瘤和恶性肿瘤两大类。 良性肿瘤生长缓慢,与周围组织界限清楚,不发生转移,对人体健康危害不大。 恶性肿瘤生长迅速,可转移到身体其他部位,还会产生有害物质,破坏正常器官结构,使机体功能失调,威胁生命。
是指细胞在致癌因素作用下,基因发生了改变,失去对其生长的正常调控,导致单克隆性异常增生而形成的新生物。这种新生物常形成局部肿块,因而得名。 根据肿瘤的生物学特性及其对机体的危害性的不同,肿瘤可分为良性肿瘤和恶性肿瘤两大类。 良性肿瘤生长缓慢,与周围组织界限清楚,不发生转移,对人体健康危害不大。 恶性肿瘤生长迅速,可转移到身体其他部位,还会产生有害物质,破坏正常器官结构,使机体功能失调,威胁生命。")
5
癌基因、抑癌基因与生长因子的关系 负调控 抑癌基因 细胞 癌基因 正调控 产物 生长因子

6
第一节 癌基因 (Oncogene) 一、概念 癌基因:细胞内控制细胞生长和分化的基因,它的结构异常或表达异常,可以引起细胞癌变。
 一、概念 癌基因:细胞内控制细胞生长和分化的基因,它的结构异常或表达异常,可以引起细胞癌变。")
7
二、癌基因理论的确立 Peyton Rous and Rous sarcoma virus

8
1966年诺贝尔奖授奖仪式上 生理学或医学奖评定委员会委员 G.克莱因教授致词
那是1910年。那时刚刚了解,每个个体细胞都通过分裂而来自其他细胞;癌细胞的分裂方式与正常细胞完全一样,不同之处仅在于它们往往肆无忌惮地侵入正常的组织屏障。 那时也刚刚认清,某些传染病是由一些小得在光学显微镜下看不见、并能通过细菌通不过的超滤器的生物所引起。这些生物称为滤过性“毒物”或病毒。 就在那时和那时前后,现代生物学尚未建立,人们还不能设想看不见的病毒与癌细胞的自行其是的生长之间会有什么关系。 大约就在这个时候,洛克菲勒研究所30岁的研究人员劳斯进行了一些实验,这些实验乍看起来有点生拉硬扯。他把一只母鸡身上的恶性结缔组织(肉瘤)制成无细胞滤液,并把它接种入健康的鸡体内。 结果是令人惊讶的,接受滤液的个体也患上了与上述母鸡完全一样的肿瘤。(说明:?)
制成无细胞滤液,并把它接种入健康的鸡体内。 结果是令人惊讶的,接受滤液的个体也患上了与上述母鸡完全一样的肿瘤。(说明:?)")
9
1932年,Shope发现,野生棉尾兔的皮肤肿瘤也可借助无细胞滤液传播。
把这些实验联系在一起,劳斯第一个想到正常细胞之变成癌细胞不一定是个突然的过程;那些原来作为整体一部分的细胞可能通过若干步骤,逐渐变成独立的自行其是的癌细胞。 劳斯把这个过程称为“肿瘤进展”,在这个过程开始时,潜在的癌细胞处于“休眠”状态。被化学因子、病毒或激素刺激唤醒之后,他们就采取一种更加无法无天的生活方式。 劳斯关于肿瘤进展的发现很快在许多实验系统内得到证实。 另一方面,他的病毒理论则备受怀疑。一般认为,病毒病都具有传染性,而癌症不能传染;这种说法如此根深蒂固,所以人们往往把各种病毒原性肿瘤解释为不可思议的例外。

10
Rous病毒是一种急转化逆转录病毒(acute transforming retrovirus,RNA病毒 )
到1950年,情况有了迅速改变。由于微生物遗传学的发展,发现某些病毒能将自己的部分遗传物质注入细胞内而不杀死细胞或抑制其增殖。这样引入的病毒物质可以整合到受体细胞的基因中,并且其行为就如同新的遗传因素一样,引起某些细胞特性的永久性改变。 重新估计了病毒概念之后,就有可能了解病毒怎么会将正常细胞循规蹈矩的行为改变为特见于癌细胞的恶性增生。 Rous病毒是一种急转化逆转录病毒(acute transforming retrovirus,RNA病毒 )
")
11
Rous 肉瘤 1966年,87岁的劳斯获得诺贝尔奖。
12
追踪研究逆转录病毒致癌的线索,Bishop等人于1980年提出了癌基因假说。
根据这一假说,引起Rous鸡肉瘤等的逆转录病毒的基因组中存在有致癌基因,即病毒癌基因,v-onc。 该癌基因来源于正常细胞基因组中的部分序列,即逆转录病毒中的致癌基因序列是在进化过程中从宿主细胞基因组中获得的。

13
反转录病毒颗粒结构 Virus-like particles assembled in vitro with purified Rous sarcoma virus Gag protein

14
* 劳氏肉瘤病毒(RSV, Rous sarcoma virus ) 基因组结构图
调节和 启动转录 LTR gag pol env src 长末端 重复序列 癌基因 正常的病毒基因 产生病毒 核心蛋白 产生逆转录 酶和整合酶 外膜蛋白 产生酪氨酸 激酶 LTR (long terminal repeat) 序列是调节区,含有反转录病毒DNA基因组表达所需要的全部调节元件 ; 在劳氏肉瘤病毒基因组中, src是额外的肿瘤式基因,它能诱发RSV感染的动物产生肉瘤
 序列是调节区,含有反转录病毒DNA基因组表达所需要的全部调节元件 ; 在劳氏肉瘤病毒基因组中, src是额外的肿瘤式基因,它能诱发RSV感染的动物产生肉瘤.")
15
LTR GAG POL ENV LTR GAG POL ENV
反转录病毒癌基因(v-onc)的起源 LTR GAG POL ENV LTR GAG POL ENV ONC
的起源. LTR. GAG. POL. ENV. LTR. GAG. POL. ENV. ONC.")
16
RNA病毒与宿主细胞基 因组整合过程示意图
感染 病毒RNA 逆转录 RNA-DNA RNA病 毒颗粒 复制 Proto-oncogenes 前病毒DNA 整合 细胞基因组 DNA 转录 病毒RNA-癌基因 宿主细胞 再感染 癌变 携带癌基因 的病毒颗粒 宿主细胞

18
二、分 类 根据来源分为病毒癌基因与细胞癌基因两类。
二、分 类 根据来源分为病毒癌基因与细胞癌基因两类。 病毒癌基因(virus oncogene,v-onc):是一类存在于肿瘤病毒中、能使靶细胞发生恶性转化的基因。 病毒癌基因不编码病毒的结构成分,对病毒复制也没有作用,但可以使细胞持续增殖。 细胞癌基因(cellular-oncogene, c-onc):存在于正常细胞基因组中的癌基因,又称原癌基因(proto-oncogene, pro-onc)。
:是一类存在于肿瘤病毒中、能使靶细胞发生恶性转化的基因。 病毒癌基因不编码病毒的结构成分,对病毒复制也没有作用,但可以使细胞持续增殖。 细胞癌基因(cellular-oncogene, c-onc):存在于正常细胞基因组中的癌基因,又称原癌基因(proto-oncogene, pro-onc)。")
19
原癌基因的特点 广泛存在于生物界中; 基因序列高度保守,为持家基因(house-keeping gene); 作用通过其产物蛋白质来体现
正常情况下表达水平很低; 被激活后,形成致癌性的细胞转化基因。

20
v-onc与c-onc 的不同 v-onc不含内含子 v-onc与c-onc的核苷酸序列不完全相同

21
断裂基因 完整的没有断裂的可读框

22
癌基因的命名 以三个斜体小写字母表示,通常源于它们的首次发现,如:
(1) src—avian Rous sarcoma virus(鸡Rous肉瘤病毒) erb—avian erythroblastosis virus(鸡成红细胞增多症病毒) sis—simian sarcoma virus(猴肉瘤病毒) ras—rat sarcomas 大鼠肉瘤 (2) 3个字母后跟一个字母或数字,说明这些基因是密切相关的成员 erbA--甲状腺激素受体的病毒类似物 erbB--表皮生长因子受体的同源类似物
 src—avian Rous sarcoma virus(鸡Rous肉瘤病毒) erb—avian erythroblastosis virus(鸡成红细胞增多症病毒) sis—simian sarcoma virus(猴肉瘤病毒) ras—rat sarcomas 大鼠肉瘤. (2) 3个字母后跟一个字母或数字,说明这些基因是密切相关的成员. erbA--甲状腺激素受体的病毒类似物. erbB--表皮生长因子受体的同源类似物.")
23
癌基因的分类 根据其基因结构和表达产物的功能特点可将原癌基因分为以下几个家族: src家族 —— 酪氨酸蛋白激酶
ras家族 —— p21蛋白,与GTP结合,GTP酶活性 myc家族 —— 核内DNA结合蛋白 sis家族 —— p28(类人血小板衍生生长因子) erb家族 —— 生长因子和蛋白激酶类 myb家族 —— 核内转录调节因子
 erb家族 —— 生长因子和蛋白激酶类. myb家族 —— 核内转录调节因子.")
25
二、癌基因的产物与功能 1、生长因子(GF):PDGF、EGF、TGF-2 2、生长因子受体 3、细胞内信号转导体 4、核内转录因子
目前认为广义的“癌基因”应当是:凡能编码生长因子、生长因子受体、细胞内生长信息传递分子,及与生长有关的转录调节因子的基因均应归属癌基因的范畴。

26
三、原癌基因激活的机制

27
(一)点突变 基因点突变是癌基因活化的主要方式。 1982年,美国 3 家研究机构(麻省理工大学、国立癌症研究所和哥伦比亚大学)几乎同时发表这样的实验结果:人膀胱癌细胞EJ株的恶性转化是由于其原癌基因 c-H-ras(356个碱基) 的点突变所致。 正常细胞H-ras碱基序列 ATG ACG GAA TAT AAG CTG GTG GTG GTG GGC GCC GGC GGT GTG 肿瘤细胞H-ras碱基序列 ATG ACG GAA TAT AAG CTG GTG GTG GTG GGC GCC GTC GGT GTG 正常p21蛋白氨基酸序列 Met Thr Glu Tyr Lys Leu Val Val Val Gly Ala Gly Ala Val 肿瘤p21蛋白氨基酸序列 Met Thr Glu Tyr Lys Leu Val Val Val Gly Ala Val Ala Val

28
突变的Ras癌基因蛋白质被锁定在激活状态 突变的Ras癌基因蛋白质被锁定在激活状态
GDP Pi GTP 无活性的Ras蛋白 内源性GTP酶活性 输入的信号刺激GDP-GTP交换 有活性的Ras蛋白 突变的Ras癌基因蛋白质被锁定在激活状态 GTP GDP 输出信号

29
Harrey和Kirfen鼠肉瘤病毒上发现,称H-ras、K-ras,人神经母细胞瘤上发现N-ras,基因突变往往发生在12、13和61密码子,以12密码子最常见。

30
突变原理: 一些化学诱变剂,如亚硝酸盐能够引起鸟嘌呤甲基化。如不能被修复,则在复制时,鸟嘌呤被腺嘌呤取代,则其 p21 蛋白的序列发生点突变,导致甘氨酸 → 天冬氨酸。 12 位 甘氨酸 天冬氨 酸 GC GC GC GC AC AT CG CG CG O6 甲基化 复制

31
(二)获得启动子与增强子 指来源于病毒等的启动子或增强子插入到细胞癌基因的附近或内部而使其开放并异常转录。
如鸡白细胞增生病毒引起的淋巴瘤,就是由于病毒的DNA序列整合到宿主细胞c-myc基因附近,成为该基因的强启动子,导致c-myc基因过强表达。

32
(avian leucosis virus,ALV)
缺乏癌基因的逆转录病毒致癌模式 鸡(禽)白细胞病毒 (avian leucosis virus,ALV) 逆转录病毒 c-myc 基因 exon-1 exon-2 exon-3 正常宿主细胞 exon-1 LTR LTR exon-2 exon-3 ALV 病毒DNA的LTR被整合到 宿主细胞c-myc癌基因附近 成为其启动子和增强子 LTR exon-2 exon-3 exon-1 ALV c-myc癌基因表达为正常的30-100倍 exon-1 LTR exon-2 exon-3 ALV
白细胞病毒. (avian leucosis virus,ALV) 逆转录病毒. c-myc 基因. exon-1. exon-2. exon-3. 正常宿主细胞. exon-1. LTR. LTR. exon-2. exon-3. ALV. 病毒DNA的LTR被整合到. 宿主细胞c-myc癌基因附近. 成为其启动子和增强子. LTR. exon-2. exon-3. exon-1. ALV. c-myc癌基因表达为正常的30-100倍. exon-1. LTR. exon-2. exon-3. ALV.")
33
(三)癌基因的甲基化程度降低 真核DNA约有5%的胞嘧啶被甲基化
癌基因的甲基化程度降低 真核DNA约有5%的胞嘧啶被甲基化")
34
DNA甲基化的生物学意义 控制基因表达 抵御外来入侵的寄生DNA
甲基化范围与基因表达程度呈反比。 DNA甲基化能关闭某些基因的活性 去甲基化则诱导了基因的重新活化和表达 抵御外来入侵的寄生DNA 近年来DNA甲基化已成为肿瘤研究的热点,甲基化状态分析可作为判断肿瘤生物学特征及临床预后的重要指标之一

35
(四)原癌基因扩增 使每个细胞中基因拷贝数增加,从而直接增加可用的转录模板数以增加基因表达。 基因扩增 蛋白质结构未变化,但总量大大提高
Cellular proto-oncogenes have been found in multiple copies in various tumors and transformed cell lines. The amplified proto-oncogene copies can occur in homogeneously staining chromosomal regions or double-minute chromosomes. The mechanism of gene amplification is not fully understood, illegitimate DNA replication occurring more than once during a single cell cycle could account for the increase of multiple segments (amplification units) of DNA from 200 to 2000 kb in size. All amplified proto-oncogenes express high levels of the corresponding RNA and protein and appear to be unrearranged. The amplified proto-oncogene is frequently tumor-specific; for example, N-myc or c-myc may be associated with the progression of neuroblastomas and small cell lung carcinoma cells. This suggests that increased expression of these proto-oncogenes plays a role in the development and progression of these tumors. 蛋白质结构未变化,但总量大大提高
 of DNA from 200 to 2000 kb in size. All amplified proto-oncogenes express high levels of the corresponding RNA and protein and appear to be unrearranged. The amplified proto-oncogene is frequently tumor-specific; for example, N-myc or c-myc may be associated with the progression of neuroblastomas and small cell lung carcinoma cells. This suggests that increased expression of these proto-oncogenes plays a role in the development and progression of these tumors. 蛋白质结构未变化,但总量大大提高.")
36
c-myc基因 c-myc基因是禽类白血病增生病毒(ALV)癌基因的同系物。
在细胞静止期,c-myc几乎不表达,但在有丝分裂原刺激下迅速表达促使细胞由G0期进入G1期,增加细胞数量。因此,c-myc原癌基因与细胞周期调控有关。 c-myc原癌基因激活的主要机制:扩增、重排和异常高表达。另外,c-myc基因低甲基化可能为其激活的另一重要机制。 1985年,Shibuya首次在部分胃癌细胞中发现c-myc基因扩增和高表达。

37
(五)基因易位或重排 由于染色体重排而导致细胞癌基因从正常位置转移到另一位置,常常是插入一启动子后而使其转录活性增加。 费城染色体。
1960年,Nowell和Hungefora,在美国费城,发现慢性粒细胞白血病的患者中,有一个很小的近端着丝粒染色体,称为费城染色体(Philadelphia chromosome, Ph) Ph染色体已被公认为慢性粒细胞性白血病的特异标记染色体。
 Ph染色体已被公认为慢性粒细胞性白血病的特异标记染色体。")
38
c-abl原癌基因的易位激活 c-abl原癌基因,经重排后插入到bcr基因的启动子后,使其转录活性增加,从而引起慢性粒细胞性白血病。
c-bcr c-abl c-abl原癌基因,经重排后插入到bcr基因的启动子后,使其转录活性增加,从而引起慢性粒细胞性白血病。

39
双星拱月 夜空现巨大笑脸 2008年12月1日,夜幕降临后,夜空中最明亮的三颗天体——月亮、金星和木星在西南方的低空齐汇人马座。
在西方神话中,金星是主管爱情和美丽的“维纳斯”女神。而木星则是“行星之王”。

40
Section 2 Anti-oncogene
第二节 抑癌基因 Section 2 Anti-oncogene

41
一、抑癌基因的基本概念 抑癌基因又称为抗癌基因(anti-oncogene)或肿瘤抑制基因,是指存在于正常细胞中,其编码产物能抑制细胞生长、增殖的一组结构基因。 抑癌基因大多编码与细胞周期调控有关的抑制蛋白,当抑癌基因缺失或突变,细胞增值失控,导致肿瘤。
或肿瘤抑制基因,是指存在于正常细胞中,其编码产物能抑制细胞生长、增殖的一组结构基因。 抑癌基因大多编码与细胞周期调控有关的抑制蛋白,当抑癌基因缺失或突变,细胞增值失控,导致肿瘤。")
42
抑癌基因与原癌基因在生物学上有以下几点差异:
①在功能上,抑癌基因起负调控作用,而原癌基因起正调控作用; ②在遗传方式上,原癌基因是显性的,而抑癌基因在细胞水平上是隐性的 抑癌基因是隐性基因,两份等位基因都突变或失活时才能使细胞转化,发现与分离难度较大

43
抑癌基因的表示方法 通常用2或3个字母编码,有时也介入蛋白质产物的大小 基因的蛋白产物的表示
Rb—retinoblastoma 视网膜母细胞瘤 Bcl-2—B-cell lymphoma B淋巴细胞瘤 p53—编码的蛋白质分子量53KD 基因的蛋白产物的表示 在其分子量前面加p,或与基因相同的代码加蛋白字样 ras产生的蛋白: Ras蛋白, p21

44
与癌基因相比,抑癌基因数量有限,说明抑癌基因比癌基因具有更普遍的作用。

45
二、抑癌基因的作用机制 (一)视网膜母细胞瘤基因 ( retinoblastoma gene,Rb)
视网膜母细胞瘤基因 ( retinoblastoma gene,Rb)")
46
视网膜母细胞瘤(retinoblastoma,Rb)
儿童期就发病的眼科恶性肿瘤,猫眼。 遗传型多为双侧发病,发病早,有家族史。 非遗传型多为单侧发病,且在2岁以后才发病。
47
1. Rb基因的结构 Rb(retinoblastoma gene)是第一个被克隆的抑癌基因。
1986年,美国的三个实验室分别独立的克隆了该基因。 Rb基因, 定位于13q14,全长约200kb,含27个外显子,26个内含子,转录的mRNA有4.7kb,编码分子量是 kD的核内磷酸化蛋白,又称p105-RB。

48
2. Rb基因的生物学活性及作用机制

49
G0 G1期 Rb蛋白 E2F DNA S期 Rb蛋白 E2F DNA mRNA (活性型) (无活性型) (转录因子,与细胞增值有关) P
周期蛋白依赖的蛋白激酶(cyclin dependence protein kinase, CDK) S期 P Rb蛋白 (无活性型) E2F DNA mRNA
 S期. P. Rb蛋白. (无活性型) E2F. DNA. mRNA.")
50
P105-Rb的作用机制(演示) G0 G1期 Rb蛋白 E-2F DNA Rb蛋白 S期 P E-2F DNA mRNA
 G0 G1期 Rb蛋白 E-2F DNA Rb蛋白 S期 P E-2F DNA mRNA")
51
RB基因的失活 某些DNA病毒癌基因表达蛋白可与RB蛋白结合形成复合物,使RB蛋白丧失抑制细胞增殖的生物特性。
如腺病毒的E1A蛋白、SV40的T抗原、人乳头瘤病毒的E7蛋白等。

52
(二) p53 基因—“基因组的卫士” 是cancer suppressive gene 一颗明珠;
Pubmed有超过4万篇论文,篇篇内容不同; 临床上, 超过50%的肿瘤其p53有异常; 曾被误认为是癌基因,直到1989年,才被证实为抑癌基因.

55
p53基因的结构 p53基因全长20kb,定位于人类染色体17p13,由11个外显子组成,编码393个氨基酸组成的53kd的核内磷酸化蛋白;
P53蛋白可分为三个结构域:N端的酸性区、中段疏水核心区与C端的碱性区,分子中有一核定位信号,所以它一般都位于核内。 酸性区 核心区 碱性区 P53蛋白 转录激活作用 结合DNA 核定位、结合DNA等

56
p53的生物学特性 p53 是细胞生长周期中的负调节因子,与细胞周期的调控、DNA修复、细胞分化、细胞凋亡等重要的生物学功能有关。

57
p53蛋白的生物学功能 ① 抑制细胞周期 ② 监测DNA损伤
p53蛋白促进p21基因表达产生分子量为21kD的蛋白质(p21WAF1/CIP1),p21是CDK的抑制剂,诱导细胞生长停滞于G1期。 ② 监测DNA损伤 DNA 损伤后,p53蛋白表达急剧上调,使细胞分裂停止于G1期,启动DNA修复系统。
,p21是CDK的抑制剂,诱导细胞生长停滞于G1期。 ② 监测DNA损伤. DNA 损伤后,p53蛋白表达急剧上调,使细胞分裂停止于G1期,启动DNA修复系统。")
58
③ 抑制DNA复制: ④ 参与程序性细胞凋亡: p53蛋白可与复制蛋白A(RPA) 结合
p53蛋白与RPA结合后可抑制RPA与DNA复制起始点的结合,阻止细胞进入S期。 p53蛋白还可通过抑制解链酶的活性,以抑制DNA复制。 ④ 参与程序性细胞凋亡: p53 蛋白使损伤细胞停留在G1期,使DNA损伤修复后再进入细胞周期。 若损伤不能修复,则启动细胞进入程序性凋亡(细胞自杀)。
。")
59
p53蛋白的作用机制(演示) 细胞自杀 复制因子A p53蛋白 DNA损伤 p53蛋白 p21基因 p53蛋白 抑 制 p21蛋白 解链酶
成功修复 p53蛋白 DNA损伤 修复失败 细胞自杀 p53蛋白 p21基因 p53蛋白 抑 制 p21蛋白 解链酶 细胞停滞于G1期

60
肿瘤中p53的突变 突变的形式是: 点突变、缺失突变、插入突变、移码突变、基因重排。 绝大多数突变集中在第5-8外显子处,即“热点突变”。

61
美国前副总统汉弗莱<Humphrey)
在1967年发现膀胱内有一肿块,病理切片未发现癌细胞 良性“慢性增生性囊肿”,未进行手术治疗。 九年后,他被诊断为患有“膀胧癌”,两年后死于该病。

62
1994年,研究者用灵敏的PCR技术对上述汉弗莱1967年的病理切片进行了p53抑癌基因检查;
这说明,在典型症状出现之前的很长时间,细胞癌变的信息已经在基因上表现出来了。

63
三、肿瘤发生是多因素、多基因和多阶段相互协调作用的癌变过程
多种遗传缺陷:点突变、易位、基因重排、移码突变、缺失突变和插入突变等 癌基因产物之间协同:核内癌基因产物与胞浆癌基因产物发生协调作用,如Myc与Ras。 癌基因与抑癌基因以及其它相关基因之间的协调

64
结直肠癌发生发展过程中的基因改变模式图 (ER Fearon和B Vogelstein , 1990年)
")
65
APC基因 APC基因,1986年由Herrera等发现,位于染色体5q21上,表达产物由2843个氨基酸残基组成。
Dihlmann等对500多个家族性多发性腺瘤病(familial adenomatous polyposis,FAP)家族进行APC基因检测,约8O个家族有APC基因突变。这些突变大多为无义突变、小插入或缺失。 因此,APC基因被认为是结直肠癌发生的早期标志之一。
家族进行APC基因检测,约8O个家族有APC基因突变。这些突变大多为无义突变、小插入或缺失。 因此,APC基因被认为是结直肠癌发生的早期标志之一。")
66
K-ras基因 K-ras基因是一种原癌基因,它的激活能导致肿瘤发生。 激活有多种形式:表达增加,基因突变,内在的GTP酶活性下降等。
超过5O%的结直肠腺瘤患者出现K-ras的过表达。 K-ras基因在结直肠癌中具有高频率的突变,并且突变出现在结直肠癌的早期,可以作为结直肠癌早期检测的标志。

67
p53基因 p53基因是重要的抑癌基因,位于染色体17pl3。
研究发现,结直肠癌患者l7号染色体短臂等位缺失,断裂点位于p53基因位点,同时发现另一等位基因上p53基因的编码区存在突变。 由于17号染色体短臂等位缺失多发生在结直肠癌患者中,而在腺瘤患者中发现较少,所以它被认为是腺瘤发展成为癌瘤的重要标志。

68
本章要点(point) 概念:癌基因,抑癌基因,病毒癌基因,细胞癌基因(或原癌基因) 病毒癌基因与原癌基因的不同 原癌基因激活的机制
点突变,获得启动子与增强子,甲基化程度降低,基因扩增,基因的易位或重排 抑癌基因 Rb基因,p53基因

69
名词解释:癌基因、抑癌基因、病毒癌基因、细胞癌基因(或原癌基因)
思考题 名词解释:癌基因、抑癌基因、病毒癌基因、细胞癌基因(或原癌基因) 试解释病毒癌基因与与对应原癌基因的不同。 试述原癌基因激活的机制。 为什么说p53是“基因卫士”?
 试解释病毒癌基因与与对应原癌基因的不同。 试述原癌基因激活的机制。 为什么说p53是 基因卫士 ?")
Similar presentations
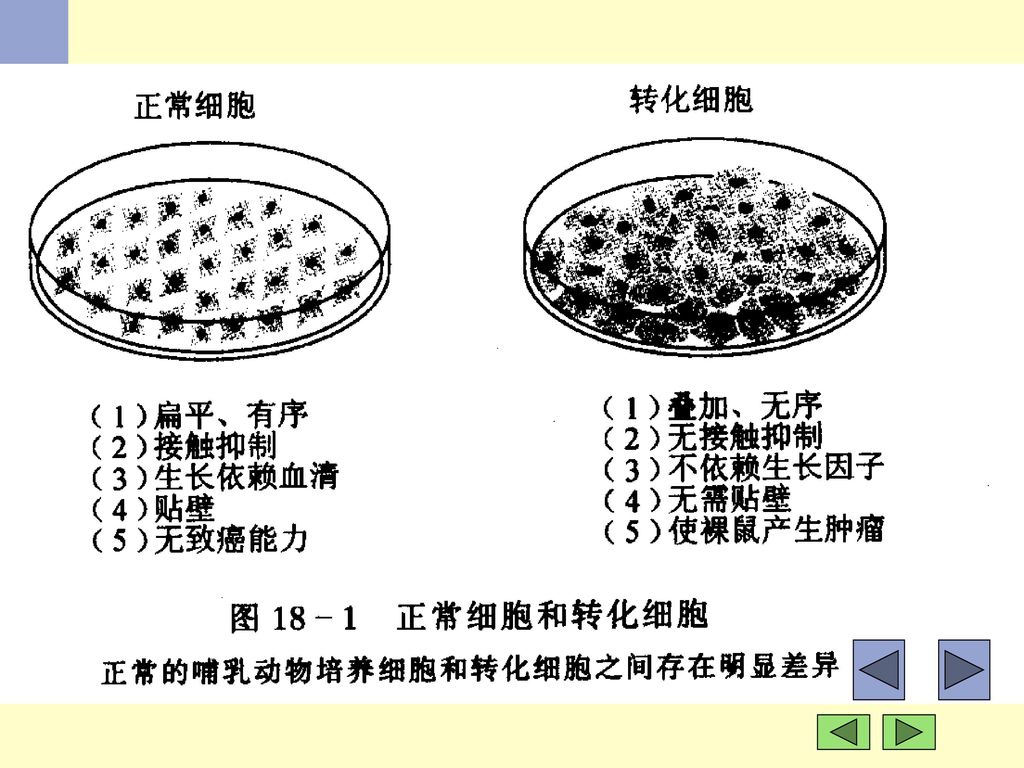



 (一)概述: ﹡ 细胞原癌基因 启动子插入 染色体移位 活化 点突变 基因扩增 癌基因 转录合成 生长因子 生长因子受体 癌基因产物(蛋白) 传导因子 核蛋白 转化蛋白 细胞恶性转化.>")

 、外环境因素 1 、化学致癌因素 已达 1000 多种 直接致 癌物 间接致 癌物 引起与机体直接 接触部位的癌肿 以前致癌物形式进 入机体,经代谢、 转化形成活化的终 致癌物.>")



 是指恶性肿瘤细胞在体内外分化诱导 剂的作用下,向正常或接近正常细胞方向分化逆转的现象。>")
癌家族 (cancer family) M 某家系中恶性肿瘤的发病率高 (约 20% )发病年龄早,通常呈 AD 遗传,其中某类肿瘤(腺癌)的发.>")
:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")
Genetics>")

— 一个特殊的“社会”(society); 细胞 — “社会”中的个体(individuals);>")

和抗癌基因(抑癌基因)的概念>")
Genetics>")
>")


是一群不受生长调控而繁殖的细胞,也称恶性肿瘤。良性肿瘤则是一群仅局限在自己的正常位置,且不侵染周围其它组织和器官的细胞。>")