Download presentation
Presentation is loading. Please wait.

2
5 抗原与获得性免疫应答

3
5-1 抗原基本概念 能够通过与 TCR 或者 BCR 特异结合而 激活 T 或者 B 淋巴细胞、诱导正性或负 性免疫应答的物质称为抗原。正性应答 导致抗体或 / 和效应 T 细胞的产生,而负 性反应则引起抗原特异性的无反应状态, 即免疫耐受 (immunological tolerance )。
。")
4
Antigen (抗原) Antigenicity (抗原性) Immunogen (免疫原) Immunogenicity (免疫原性) Toleragen (耐受原) Allergen (变应原)
 Antigenicity (抗原性) Immunogen (免疫原) Immunogenicity (免疫原性) Toleragen (耐受原) Allergen (变应原)")
5
大于 Ab 或者 TCR 的抗原结合部位 无机物不能成为抗原; 分子量太小(如对氨基苯甲酸, DNP )的有机物不能成为抗原; 蛋白质是最主要、最常见的抗原; 多糖、脂肪、核苷酸等有机大分子能被 B 细胞和 -T 细胞所识别。 抗原的基本理化特性 ——
的有机物不能成为抗原; 蛋白质是最主要、最常见的抗原; 多糖、脂肪、核苷酸等有机大分子能被 B 细胞和 -T 细胞所识别。 抗原的基本理化特性 ——")
6
抗原的免疫原性( immunogenecity ) 分子量在 5 kDa 以上含有蛋白质成分的有机大分子,纯化的多糖、脂 类及核苷酸等物质难以单独诱导 T 和 B 淋巴细胞活化。 分子结构越复杂(含有多个 T 和 B 细胞表位)免疫原性越强。 抗原与被免疫宿主之间的遗传关系越远,免疫原性越强。 皮内、皮下、肌肉、静脉和腹腔等途径诱导正性免疫应答。 经口腔和消化道摄入的抗原更倾向于诱导免疫耐受(负性免疫应答)。
 分子量在 5 kDa 以上含有蛋白质成分的有机大分子,纯化的多糖、脂 类及核苷酸等物质难以单独诱导 T 和 B 淋巴细胞活化。 分子结构越复杂(含有多个 T 和 B 细胞表位)免疫原性越强。 抗原与被免疫宿主之间的遗传关系越远,免疫原性越强。 皮内、皮下、肌肉、静脉和腹腔等途径诱导正性免疫应答。 经口腔和消化道摄入的抗原更倾向于诱导免疫耐受(负性免疫应答)。")
7
1 无应答 -- 抗原浓度太低或者宿主已经处于耐受状态 2 抗原特异性体液和细胞免疫应答(正性应答) 宿主此后的一段时间里对该抗原处于免疫状态 3 超敏反应 抗原特异性免疫应答伴有较强的炎症反应或损伤 4 诱导免疫耐受(负性应答) 宿主在此后的一段时间里对该抗原处于无反应状态 外来抗原进入体内可能产生四种不同的结果:
 宿主此后的一段时间里对该抗原处于免疫状态 3 超敏反应 抗原特异性免疫应答伴有较强的炎症反应或损伤 4 诱导免疫耐受(负性应答) 宿主在此后的一段时间里对该抗原处于无反应状态 外来抗原进入体内可能产生四种不同的结果:")
8
诱导免疫耐受的耐受原 100 抗原浓度 耐受

9
抗原剂量与免疫应答的关系 一般情况下外来抗原进入体内会引起体 液和细胞免疫应答,但是高剂量或者低剂 量反复静脉注射可溶性抗原则能够在 4 - 5 天内造成抗原特异性 B 细胞无反应状态。用 大剂量抗原诱导的免疫耐受状态要靠高浓 度抗原维持。如果在 B 细胞进入活性封闭 24 小时之内撤去耐受抗原并辅以 T 细胞帮助, B 细胞能够得到挽救。

10
1 抗原本身的特性 —— 非聚合物容易诱导耐受; 佐剂诱导正性免疫反应; 右旋多肽诱导耐受而左旋的易诱导免疫反应。 2 宿主年龄 —— 年令越小越容易诱导耐受。 3 免疫抑制药剂与抗体 —— 辅助诱导抗原特异的免疫耐受。 4 诱导途径和方式 —— 静脉、口服、低剂量的抗原的反复注射。 诱导免疫耐受的一般规律:

11
抗原抗原 B 细 胞 表 位 抗体 CDR3 表位 T 和 B 细胞表位( epitope ): 抗原分子中被 TCR 或者 BCR 直接 识别的部分构成 T 或者 B 细胞表位。 B 细胞表位: 位于抗原分子的表面 具有构象依赖性
: 抗原分子中被 TCR 或者 BCR 直接 识别的部分构成 T 或者 B 细胞表位。 B 细胞表位: 位于抗原分子的表面 具有构象依赖性")
12
B 细胞表位 (B cell epitope) 抗原表面能够与抗体结合的最小单位即 B 细胞表位,具有构象依赖性。抗原表 位与抗体分子的抗原结合部位在形状上 互补才能保证抗原与抗体的紧密结合。
 抗原表面能够与抗体结合的最小单位即 B 细胞表位,具有构象依赖性。抗原表 位与抗体分子的抗原结合部位在形状上 互补才能保证抗原与抗体的紧密结合。")
13
Ep1Ep2Ep3 T 细胞表位 ( T Cell epitope ) 能够与 MHC 并被 TCR 识别的肽段 与蛋白质分子构象无关 与肽段在蛋白质分子中的位置无关 主要决定于肽段的氨基酸序列 与宿主表达的 MHC 分子有关
 能够与 MHC 并被 TCR 识别的肽段 与蛋白质分子构象无关 与肽段在蛋白质分子中的位置无关 主要决定于肽段的氨基酸序列 与宿主表达的 MHC 分子有关")
14
流感病毒血凝素分子流感病毒血凝素分子

15
TD ( T-dependent )抗原 和 TI ( T-independent )抗原 天然抗原 ( Natural ) 与 人工抗原 自身抗原( auto-, self- ) 与 外来抗原( foreign ) 抗原的分类
抗原 和 TI ( T-independent )抗原 天然抗原 ( Natural ) 与 人工抗原 自身抗原( auto-, self- ) 与 外来抗原( foreign ) 抗原的分类")
16
1 2 1 2 CD40/CD40L B 细胞 Th 细胞 TI-1 抗原 TD 抗原 TI 与 TD 抗 原 CD4

18
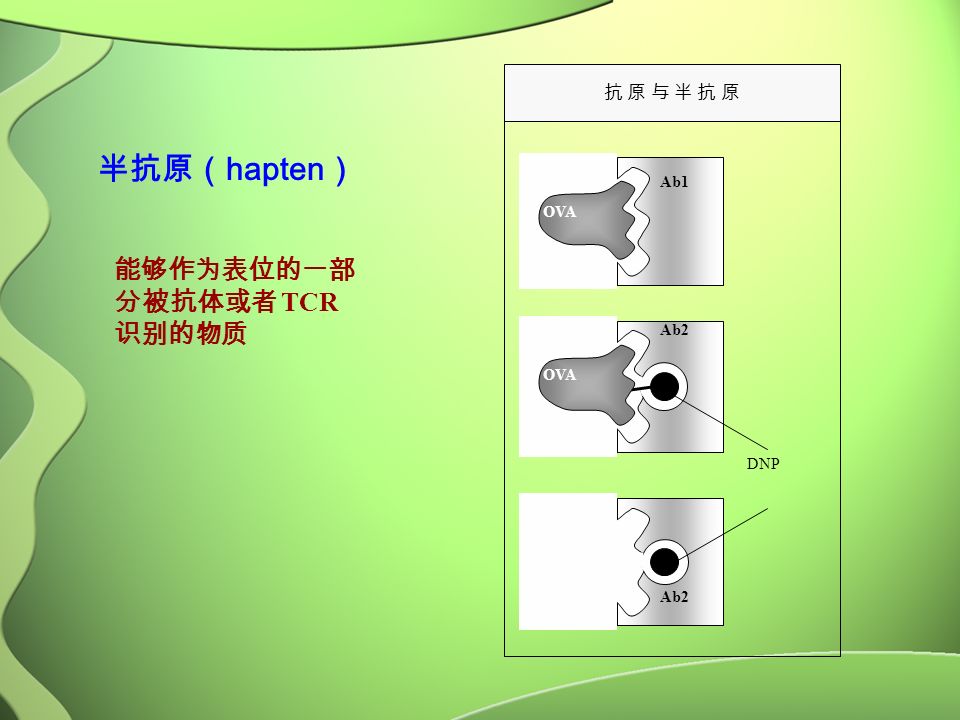
OVA 抗 原 与 半 抗 原抗 原 与 半 抗 原 Ab2 Ab1 DNP OVA 半抗原( hapten ) 能够作为表位的一部 分被抗体或者 TCR 识别的物质
 能够作为表位的一部 分被抗体或者 TCR 识别的物质")
19
半抗原 半抗原 DNP 本身没有诱导免疫应答的能 力。但是用与 OVA 偶联的 DNP 免疫小鼠, 所得到的抗体( Ab2 )能够在体外与半抗 原特异地结合。在这种情况下 DNP 成为 B 细胞表位的一个部分。
能够在体外与半抗 原特异地结合。在这种情况下 DNP 成为 B 细胞表位的一个部分。")
20
2m TCR- chain TCR- chain Homocysteine HLA-A68 Peptide Interaction between TCR and homocysteine presented by HLA-A68 C

21
T 细胞半抗原 —— 能够修饰 MHC/ 抗原肽的分子可能作为半 抗原被 TCR 识别。

22
超抗原 TCRV 抗原肽 MHC-II T 细胞 APC 超抗原

23
超抗原 —— 超抗原能与 MHC-II 分子及 TCR 链结 合,从而造成 T 细胞的非特异性活化。

24
免疫佐剂

25
5-2 获得性免疫应答的基本特点 1 One cell , one receptor 2 Clonal selection and expansion 3 Clonal deletion 4 Lymphoid organs 5 Antigen processing and presentation ( MHC )
")
26
MHC 分子与抗原识别 淋巴细胞如何识别天文数字的抗原 次级淋巴器官与免疫识别 抗原特异性淋巴细胞的克隆扩增与选择 免疫耐受 -- Clonal deletion

27
5-3 天然免疫应答和获得性免疫应答之间的关系 获得性免疫应答的启动 巨噬细胞和树突细胞的活化与否对启动获得性免疫应答起着 “ 控制阀 ” 的作用。 如果没有巨噬细胞和树突细胞的先期反应,获得性免疫应答也就无从谈起。 外来抗原也可以在没有 APC 的 “ 陪伴 ” 下进入引流淋巴结,但它们必须首先被补体 片段致敏。

28
抗体是天然免疫系统的 “ 眼睛 ” 抗体分子与抗原结合后活化补体,使其更为准确、集中、有效地杀伤靶细胞。 介导 NK 和巨噬细胞的抗体依赖的细胞毒效应( ADCC )。 T 细胞与天然免疫细胞之间的协同作用 细胞免疫应答所产生的效应 T 细胞通过分泌 IFN- 和 IL-18 等细胞因子促进巨噬细 胞的活化,使之更加有效地吞噬、消化外来抗原或者微生物。 巨噬细胞分泌 IL-1 和 IL-12 等细胞因子,诱导 T 细胞的分化。 天然免疫与获得性免疫的时间互补 获得性免疫系统在受到外来抗原刺激之后一般需要 5~7 天方能发挥效应,而天然 免疫系统是抗感染免疫过程中的 “ 先头部队 ” ,在微生物进入机体几分钟之内就会 开始应答。起到限制微生物在体内迅速扩散的作用。
。 T 细胞与天然免疫细胞之间的协同作用 细胞免疫应答所产生的效应 T 细胞通过分泌 IFN- 和 IL-18 等细胞因子促进巨噬细 胞的活化,使之更加有效地吞噬、消化外来抗原或者微生物。 巨噬细胞分泌 IL-1 和 IL-12 等细胞因子,诱导 T 细胞的分化。 天然免疫与获得性免疫的时间互补 获得性免疫系统在受到外来抗原刺激之后一般需要 5~7 天方能发挥效应,而天然 免疫系统是抗感染免疫过程中的 先头部队 ,在微生物进入机体几分钟之内就会 开始应答。起到限制微生物在体内迅速扩散的作用。")
Similar presentations

人体免疫系统在维持稳态中的作用 ( 5 )艾滋病的流行和预防.>")









>")

>")






