Download presentation
1
(RNA interference,RNAi)
007小组组长:方刚 组员:张科强 范宇巧 田甜 肖鹏 车斌

2
Contents RNAi的发现简史 1 RNAi的相关知识 2 RNAi的分子机制 3 RNAi的应用前景 4 RNAi的实验方法 5

3
2006年10月2日,瑞典卡罗林斯卡医学院宣布将2006年度诺贝尔生理学或医学奖授予两名美国科学家安德鲁·法尔和克雷格·梅洛,以表彰他们发现了“RNA(核糖核酸)干扰”机制。
法尔目前任职于美国麻省理工学院,梅洛目前在美国哈佛大学工作。他们将分享1000万瑞典克朗(约合140万美元)的奖金。
的奖金。")
4
梅洛(Craig Mello)生于1960年,是被恐龙骨引入科学世界的。梅洛童年时经常跟着父亲在美国西部寻找化石。从那时起,他就迷上了远古时代、地球历史和人类生命的起源等问题。高中时代,梅洛的兴趣逐渐转移到了基因工程方面。 法尔(Andrew Fire)1959年出生在美国加利福尼亚州,本科在加利福尼亚大学伯克利分校主修数学,仅用3年时间就拿到学位。与梅洛类似,他逐渐对涉及生命奥秘的遗传学产生兴趣,并将其作为自己终身的学术追求。
1959年出生在美国加利福尼亚州,本科在加利福尼亚大学伯克利分校主修数学,仅用3年时间就拿到学位。与梅洛类似,他逐渐对涉及生命奥秘的遗传学产生兴趣,并将其作为自己终身的学术追求。")
5
下面就让我们来看看RNA干扰的发现简史。
06年获奖时,梅洛46岁,法尔47岁,在历代诺贝尔奖获得者中,是年龄偏低的,而且他们从科学发现到获奖还不足10年的时间,这在诺贝尔奖历史上还属罕见。诺贝尔生理学或医学奖评委在现场解析今年的获奖成果时说:这是诺贝尔奖首次不发给一个拥有答案的研究,相反,RNA干扰机制为我们提出的是更多需要解答的问题,为基因技术研究提供了令人兴奋的可能性。 这样一个让诺贝尔奖都为之破例的伟大发现,究竟经历了哪些历程,就这么快就被科学界承认了呢? 下面就让我们来看看RNA干扰的发现简史。

6
一、RNA干扰的发现简史 90年代初,美国和荷兰的两个转基因植物实验组Rich Jorgensen和同事,在对矮牵牛(petunias)进行的研究中有个奇怪的发现:将一个能产生色素的基因置于一个强启动子后,导入矮脚牵牛中,试图加深花朵的紫颜色,结果没看到期待中的深紫色花朵,多数花成了花斑的甚至白的。也就是说转基因的植株不仅没有新基因表达,反而使原有的色素基因也受到了抑制,Jorgensen将这种现象命名为共抑制(cosuppression) 94年Cogoni等证明真菌中亦有类似现象,此称为基因压制(quelling)。
进行的研究中有个奇怪的发现:将一个能产生色素的基因置于一个强启动子后,导入矮脚牵牛中,试图加深花朵的紫颜色,结果没看到期待中的深紫色花朵,多数花成了花斑的甚至白的。也就是说转基因的植株不仅没有新基因表达,反而使原有的色素基因也受到了抑制,Jorgensen将这种现象命名为共抑制(cosuppression) 94年Cogoni等证明真菌中亦有类似现象,此称为基因压制(quelling)。")
7
预测 Rich Jorgensen 这足以说明,植物领域科学家们在RNAi发现和机制阐述方面做出的关键性作用 试验 野生型
试验 预测 野生型 06年诺贝尔生理学或医学奖颁布后 来自英国剑桥大学生物技术学院,以及比利时法兰德斯大学校际生物科技研究所的三名科学家向英国著名的《Nature》杂志寄出的一封信中,表示了他们对于此次RNAi获得诺贝尔奖的遗憾之处——植物RNAi研究领域科学家没有能包括在这一奖项中 这足以说明,植物领域科学家们在RNAi发现和机制阐述方面做出的关键性作用 Rich Jorgensen

8
一、RNA干扰的发现简史 1995年,康乃尔大学的Su Guo博士在试图阻断秀丽新小杆线虫(C.elegans)的par-1基因时,发现了一个意想不到的现象。她们本想利用反义RNA技术特异性地阻断上述基因的表达,而同时在对照实验中给线虫注射正义RNA以期观察到基因表达的增强,但得到的结果是二者都同样地切断了par-1基因的表达途径。这是与传统上对反义RNA技术的解释正好相反。该研究小组一直未能给这个意外以合理解释。 论文: Guo S, Kemphues KJ, Par L. Cell, 1995,81:
的par-1基因时,发现了一个意想不到的现象。她们本想利用反义RNA技术特异性地阻断上述基因的表达,而同时在对照实验中给线虫注射正义RNA以期观察到基因表达的增强,但得到的结果是二者都同样地切断了par-1基因的表达途径。这是与传统上对反义RNA技术的解释正好相反。该研究小组一直未能给这个意外以合理解释。 论文: Guo S, Kemphues KJ, Par L. Cell, 1995,81:")
9
一、RNA干扰的发现简史 1998年,华盛顿卡耐基研究院的Fire和麻省大学医学院的Mello首次在秀丽新小杆线虫中证明上述现象属于转录后水平的基因沉默。他们发现Su Guo博士遇到的正义RNA抑制基因表达的现象,以及过去的反义RNA技术对基因表达的阻断,都是由于体外转录所得RNA中污染了微量双链RNA而引起。 当他们将体外转录得到的单链RNA纯化后注射线虫时发现,基因抑制效应变得十分微弱,而经过纯化的双链RNA却正好相反,能够高效特异性阻断相应基因的表达,其抑制基因表达的效率比纯化后的反义RNA至少高2个数量级。该小组将这一现象称为RNA干扰。 论文: Fire A, Xu S, Montgomery ME, et al. Nature. 1998,391:

10
一、RNA干扰的发现简史 在1999年短短的一年间,发现RNA干扰现象广泛存在于从植物、真菌、线虫、昆虫、蛙类、鸟类、大鼠、小鼠、猴一直到人类的几乎所有的真核生物中细胞。2000年,又先后发现小鼠早期胚胎中和大肠杆菌中也存在RNA干扰现象。 2000年提出RNAi作用机制模型。 论文: Hannond SM et al. Nature, 2000, 404(6775): Zamore PD. Cell, 2000,101(1):25-33 2001年,RNAi技术成功诱导培养的哺乳动物细胞基因沉默现象。Nature,2001,411(6836):494-498
: Zamore PD. Cell, 2000,101(1): 年,RNAi技术成功诱导培养的哺乳动物细胞基因沉默现象。Nature,2001,411(6836):494-498.")
11
一、RNA干扰的发现简史 此后,RNA干扰技术迅速兴起并掀起了一股生物技术热潮,在2000~2003年间,连续4年被列入世界十大科技进步之一。Sciencee曾将RNAi评为2002年十大科技进步之首。 麻省理工大学的生物学家,诺贝尔奖获得者菲利普 夏普认为,RNAi技术将成为十年来甚至几十年来最重要和激动人心的科学突破

12
线虫体内的RNAi 线虫中RNAi效应的检测
(左面)两条线虫表现为绿色荧光蛋白(GFP)表达类型品系,该品系含有一个普遍性表达的GFP报告基因重组质粒(带有核内定位信号位点)。 (右面)喂食表达针对GFP的dsRNA的细菌后,整个虫体的GFP信号消失。
两条线虫表现为绿色荧光蛋白(GFP)表达类型品系,该品系含有一个普遍性表达的GFP报告基因重组质粒(带有核内定位信号位点)。 (右面)喂食表达针对GFP的dsRNA的细菌后,整个虫体的GFP信号消失。")
13
RNAi的定义 目前对RNAi (RNA interference)的定义有很多种,不同的资料对其定义的侧重点也不尽相同
① RNAi是由dsRNA介导的由特定酶参与的 特异性基因沉默现象,它在转录水平、转录后水 平和翻译水平上阻断基因的表达。 ② RNAi是有dsRNA参与指导的,以外源和内源mRNA 为降解目标的转基因沉默现象。具有核苷酸序列特异性 的自我防御机制,是一种当外源基因导入或病毒入侵后, 细胞中与转基因或入侵病毒RNA同源的基因发生共同基 因沉默的现象。

14
RNAi的定义 如果将其作为一门生物技术,则定义为:
③ RNAi是将与靶基因的mRNA 同源互补的双链RNA(dsRNA ) 导入细胞,能特异性地降解该mRNA ,从而产生相应的功能表型缺失, 属于转录后水平的基因沉默(post - transcriptional gene silence , PTGS)。 ① RNAi 是指通过反义RNA与正链RNA 形成双链RNA 特异性地抑制靶基因的现象,它通过人为地引入与内源靶基因具有相同序列的双链RNA(有义RNA 和反义RNA) ,从而诱导内源靶基因的mRNA 降解,达到阻止基因表达的目的。 ② RNAi是指体外人工合成的或体内的双链RNA(dsRNA)在细胞内特异性的将与之同源的 mRNA降解成21nt~23nt 的小片段,使相应的基因沉默。
 导入细胞,能特异性地降解该mRNA ,从而产生相应的功能表型缺失, 属于转录后水平的基因沉默(post - transcriptional gene silence , PTGS)。 ① RNAi 是指通过反义RNA与正链RNA 形成双链RNA 特异性地抑制靶基因的现象,它通过人为地引入与内源靶基因具有相同序列的双链RNA(有义RNA 和反义RNA) ,从而诱导内源靶基因的mRNA 降解,达到阻止基因表达的目的。 ② RNAi是指体外人工合成的或体内的双链RNA(dsRNA)在细胞内特异性的将与之同源的 mRNA降解成21nt~23nt 的小片段,使相应的基因沉默。")
15
我们不惜花费这样的篇幅给同学们介绍RNAi的发现史,一来是希望同学们能了解这项伟大发现的来龙去脉,知道那些曾为此做出过贡献的科学家,二来也是希望能引起同学们对RNAi的兴趣。
接下来,我们将为同学们介绍的是RNAi的相关知识,例如前面所提到过的反义RNA和基因沉默等。这是同学们可能不太了解,但是对于理解RNAi的分子机制又必不可少的东西。

16
基因沉默 RNAi的 相关知识 反义RNA DICER酶 SiRNA MicroRNA

17
基因沉默(gene silencing) 基因沉默是指生物体中特定基因由于种种原因不表达。 两种方式:
TGS (transcriptional gene silencing) PTGS (post-transcriptional gene silencing) 在这两种水平上引起的基因沉默都与基因的同源性有关,称为同源依赖性的基因沉默。 (homology-dependent gene silencing,HDGS)。
 PTGS (post-transcriptional gene silencing) 在这两种水平上引起的基因沉默都与基因的同源性有关,称为同源依赖性的基因沉默。 (homology-dependent gene silencing,HDGS)。")
18
基因沉默(gene silencing))
一方面,基因沉默是遗传修饰生物(genetically modified organisms)实用化和商品化的巨大障碍。 另一方面,基因沉默是植物抗病毒的一个本能反应,为用抗病毒基因植物工程育种提供了具有较大潜在实用价值的策略——RNA介导的病毒抗性。
实用化和商品化的巨大障碍。 另一方面,基因沉默是植物抗病毒的一个本能反应,为用抗病毒基因植物工程育种提供了具有较大潜在实用价值的策略——RNA介导的病毒抗性。")
19
反义RNA 反义RNA是指与mRNA互补的RNA分子(也包括与其它RNA互补的RNA分子)
由于核糖体不能翻译双链的RNA,所以反义RNA与mRNA特异性的互补结合, 即抑制了该mRNA的翻译。

20
细胞中反义RNA的来源有两种途径∶ 第一是反向转录的产物,在多数情况下, 反义RNA是特定靶基因互补链反向转录产物, 即产生mRNA和反义RNA的DNA是同一区段的互补链。 第二种来源是不同基因产物,如OMPF基因是大肠杆菌的膜蛋白基因,与透性有关,其反义基因MICFZE则为另一基因。

21
反义RNA的分类和作用机制 Ⅰ类:这类反义RNA直接作用于其靶mRNA的编码区,引起翻译的直接抑制(ⅠA类)或与靶mRNA结合后引起该双链RNA分子对RNA酶Ⅲ的敏感性增加,使其降解(ⅠB类)。 Ⅱ类:这类反义RNA与mRNA的SD序列的上游非编码区结合,从而抑制靶mRNA的翻译功能。其作用机制尚不完全清楚 Ⅲ类:这类反义RNA可直接抑制靶mRNA的转录。

22
dicer Dicer是RNaseIII家族中的一员,是特异识别双链RNA的核酸内切酶

23
四个结构域:Argonaute家族的PAZ结构域,III型RNA酶活性区域,dsRNA结合区域以及DEAH/DEXHRNA解旋酶活性区

24
Small interfering RNA (siRNA)
long dsRNA siRNAs 一种小RNA分子(~21-25核苷酸),由Dicer加工而成。SiRNA是siRISC的主要成员,激发与之互补的目标mRNA的沉默,是 RNAi的主要效应分子
,由Dicer加工而成。SiRNA是siRISC的主要成员,激发与之互补的目标mRNA的沉默,是 RNAi的主要效应分子.")
25
Small interfering RNA (siRNA):
2. 依赖Dicer酶的加工,是Dicer的产物,所以具有Dicer产物的特点。 3. 生成需要Argonaute家族蛋白存在。 4. 是RISC组分。 5. siRNA合成是由双链的RNA或RNA前体形成的。 6. siRNA是人工体外合成的,通过转染进入人体内,是RNA干涉的中间产物。 7. 结构上, siRNA是双链RNA。 8. 在Dicer酶的加工过程中, siRNA对称地来源于双链RNA的前体的两侧臂。 9. 在作用位置上, siRNA可作用于mRNA的任何部位。 10. 在作用方式上, siRNA只能导致靶标基因的降解,即为转录水平后调控。 11. siRNA不参与生物生长,是RNAi的产物,原始作用是抑制转座子活性和病毒感染。

26
走进MicroRNA的大世界 MicroRNA也可以写做miRNA ,是一种21-25nt长的单链小分子RNA。它广泛存在于真核生物中,是一组不编码蛋白质的短序列RNA。成熟的miRNA,5′端有一个磷酸基团,3′端为羟基。

27
MicroRNAs: Expanding Family of ‘RiboRegulators’
其碱基分子能够与那些和它的序列互补的mRNA分子相结合,有时候甚至可以与特定的DNA片断结合。这种结合的结果就是导致基因的沉默。这种方式是身体调节基因表达的一个重要策略。据推测,miRNA调节着人类三分之一的基因。

28
MiRNA的作用方式 以线虫lin-4为代表,作用时与靶标基因不完全互补结合,进而抑制翻译,但是不影响mRNA的稳定性,这种mRNA是目前发现的最多的种类。 以拟南芥miR-171为代表,作用时与靶标基因完全互补结合,作用方式与siRNA相似,最后切割靶mRNA。 具有以上两种作用模式,当与靶基因完全结合时,切割mRNA,不完全互补结合时,起调节作用。

29
miRNA与近亲siRNA的差异 在描述两者的差异之前,有必要先说一说它们的共同点:
2. 它们都是Dicer酶的产物; 3. 它们在起干扰、调节作用时都会和RISC诱导的沉默 复合体(RNA-induced silencing complex,RISC)结合; 4. 它们都可以在转录后和翻译水平干扰以抑制靶标基因的翻译;
结合; 4. 它们都可以在转录后和翻译水平干扰以抑制靶标基因的翻译;")
31
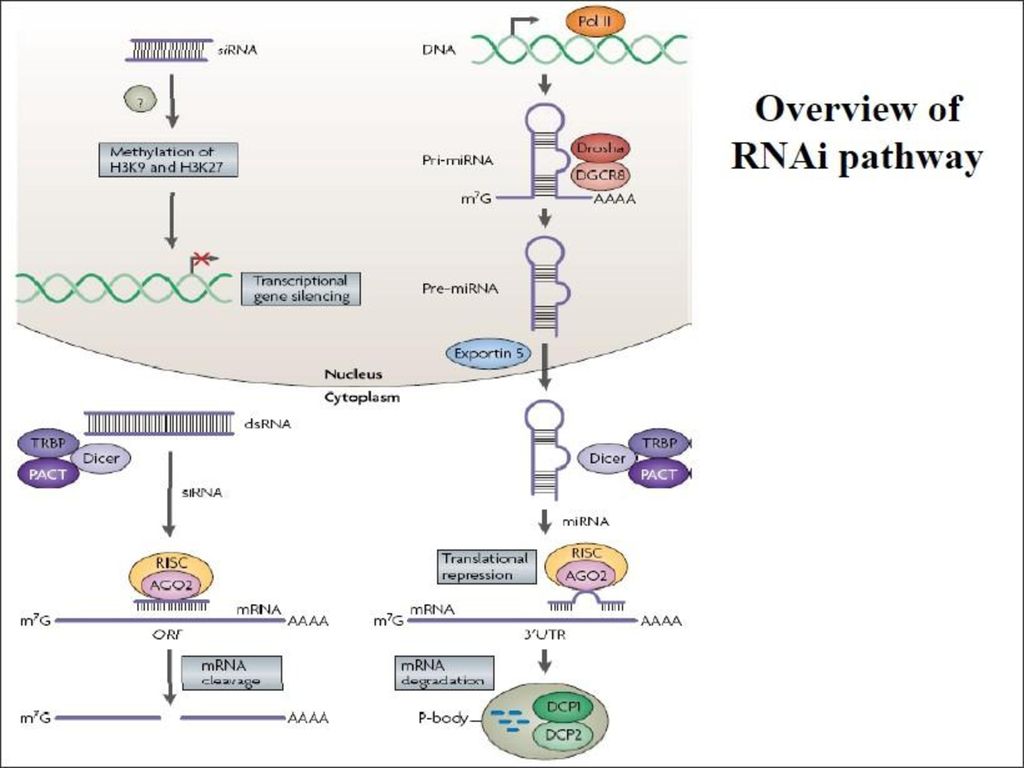
RNA干扰的分子机制: 关于这个问题,目前阐明的并不是很清楚,其中很多问题都还没有解决,但是大体模型已经建立。

32
RNAi works postranscriptionally……..
How does RNAi work? RNAi works postranscriptionally…….. in key two steps!

33
processing the dsRNA into 21-23 nt fragments
step one: processing the dsRNA into nt fragments 34 27 21 20 16 short-interfering RNA 第一步(起始阶段)是较长ds RNA在ATP参与下被RNaseⅢ样的特异核酸酶切割加工成21~23nt的由正义和反义链组成的小干扰RNA(small interfering RNA,siRNA)。 Tuschl, 2001
是较长ds RNA在ATP参与下被RNaseⅢ样的特异核酸酶切割加工成21~23nt的由正义和反义链组成的小干扰RNA(small interfering RNA,siRNA)。 Tuschl,")
34
siRNAs have a defined structure
19 nt duplex 2 nt 3’ overhangs siRNA的两条单链末端为5’-磷酸和3’-羟基,且3’端均有2-3个突出的核苷酸。

35
RNAi silencing complex
may be associated with translating ribosomes active RNAse enzyme not yet identified may participate in endogenous pathways that silence genes via translational repression 第二步(效应阶段)是siRNA 在ATP参与下被RNA解旋酶解旋成单链,并由其中反义链指导形成RNA诱导的沉默复合体(RNA-induced silencing complex,RISC)。
是siRNA 在ATP参与下被RNA解旋酶解旋成单链,并由其中反义链指导形成RNA诱导的沉默复合体(RNA-induced silencing complex,RISC)。")
36
step two: the antisense strand of the siRNA guides cleavage
活化的RISC在单链siRNA引导下识别互补的mRNA,并在RISC中的核酸内切酶作用下从siRNA引导链中心所对应的靶基因位置切割靶mRNA,最后可能再被核酸外切酶进一步降解,从而干扰基因表达。 Tuschl, 2002

37
扩增和扩散阶段 siRNA不仅可引导RISC切割靶RNA,而且可作为引物在RNA依赖的RNA聚合酶(RdRP)作用下以靶mRNA为模板合成新的dsRNA。 新合成的长链dsRNA同样可被dicer切割、降解而生成大量的次级siRNA。次级siRNA又可进入合成-切割的循环过程,进一步放大RNAi作用。 Gisela Storz, Science, 296(5571): , 2002.
: ,")
39
二、RNA干扰的机制 RNAi干扰现象具有以下几个重要的特征: ①RNAi是转录后水平的基因沉默机制; ②RNAi具有很高的特异性;
③只有dsRNA才能诱导产生RNAi; ④只有针对编码区的dsRNA才能产生有效的和特异性的干扰; ⑤注射同源dsRNA可以引起内源性mRNA特异性的降解; ⑥RNAi抑制基因表达具有很高的效率,可达到缺失突变体表型的程度,而且相对很少量的dsRNA分子就能完全抑制相应基因的表达,这是通过催化放大的方式进行的;

40
二、RNA干扰的机制 RNAi干扰现象具有以下几个重要的特征:
⑦RNAi抑制基因表达的效应可以长距离传递和维持信号甚至传播至整个有机体以及可遗传给F1,但F2往往恢复为野生型。 ⑧dsRNA不得短于21个碱基,大于30 bp的dsRNA不能在哺乳动物中诱导特异的RNA干扰,而是细胞非特异性和全面的基因表达受抑和凋亡; ⑨ RNAi作用迅速,mRNA快速降解; ⑩ RNAi效应的依赖性,只有连续产生dsRNA才能产生长期效应,否则只产生短暂的沉默反应。

41
cell death Mammals exhibit potent responses to dsRNA P PKR dsRNA
interferon production eiF2a P cell death apoptosis Blockage of protein synthesis dsRNA 进入哺乳动物成体细胞后,细胞内病毒防御 机制被激活。细胞内干扰素产生增加,蛋白激酶PKR 激活,使转录因子E2F被抑制,非特异性的阻断基因的 转录,并诱导细胞凋亡。

42
siRNAs are not recognized
smaller RNAs can escape the PKR pathway recall that siRNAs are intermediate effectors In the RNAi pathway siRNAs are not recognized by the PKR!

43
RNAi调控染色质甲基化 RNA干扰的一部分作用机理:由于只有在DNA未折叠的情况下基因才会表现,因此RNAi凭借甲基化等修饰组蛋白而影响局部DNA的折叠,从调控大区域的基因表现,在发育及分化的过程中扮演了极为重要的地位。而且RNAi所诱发的染色质修饰需要RNA polymeraseII的参与,而也就说明了这种凭借甲基化染色质来抑制基因表达的过程,可能也是需要先转录成RNA,然后制造siRNA以进行染色质的修饰,抑制基因的表达。

44
siRNA and Silent Chromatin - Model
RNA homologous to centromeric repeats are processed – siRNAs siRNAs may recruit Clr4 histone H3 methylase result in meth. of H3 Lys9 Swi6 binds chromatin Gene silencing

46
总之,作为一门新兴的技术,RNA干扰中还有很多的细节没有弄清楚,比如RNA干扰的所有中介物是什么,不同RISC是如何形成的。但是这并不妨碍我们对于它的应用前景的展望。

47
RNA干扰的应用及前景 1、基因功能的研究 2、病毒性疾病的治疗 3、肿瘤的治疗 4、药物的研发

48
1.基因功能研究 由于RNAi具有高度的序列专一性和有效的干扰活力,可以特异地使特定基因沉默,获得功能丧失或降低突变,因此可以作为功能基因组学的一种强有力的研究工具。已有研究表明RNAi能够在哺乳动物中抑制特定基因的表达,制作多种表型,而且抑制基因表达的时间可以控制在发育的任何阶段,产生类似基因敲除的效应。与传统的基因敲除技术相比,这一技术具有投入少,周期短,操作简单等优势,近来RNAi成功用于构建转基因动物模型的报道日益增多,标志着RNAi将成为研究基因功能不可或缺的工具。

49
Jacque et al 针对HIV-1复制的早 期和晚期, 设计针对HIV-1基因组不同区域的
2.病毒性疾病的治疗——艾滋病 Jacque et al 针对HIV-1复制的早 期和晚期, 设计针对HIV-1基因组不同区域的 siRNAs, 并转染人类细胞系、原始淋巴细胞及 CD4+HeLa细胞, 结果证明RNAi能够降低HIV-1 基因组的RNA. 《Nature 》2002

50
Introduced 22 nucleotide synthetic siRNAs (complementary to HIV target +/- GFP) into human cell lines/ primary lymphocytes RESULTS: DO NOT ACTIVATE PKR PATHWAY and siRNAs SPECIFICALLY DEGRADE HIV-1 mRNA, dsRNA-activated protein kinase

51
2.病毒性疾病的治疗——HBV Shlomai et al 针对HBV的核心抗原开放阅读框和X蛋白开放阅读框设计了两个siRNA片段, 有效地抑制了Huh-7细胞系中HBV的复制. 《Hepatology 2003》

52
2.病毒性疾病的治疗_——HCV Kapadia et al 利用RNAi技术, 对HCV进行研究, 用针对HCV的2个RNAi与表达HCV的质粒共转染人的肝癌细胞株Huh-7细胞, 2 d后, Northern杂交检测HCV的RNA含量降低, 证明HCV特异的siRNA抑制了病毒的复制. Proc Natl Acad Sci USA 2003

53
科学家已经用RNAi研究过如:人流感病毒、柯萨奇病毒B3 (CVB3)、人多瘤病毒JCV等病毒。
2.病毒性疾病的治疗_——其它病毒 科学家已经用RNAi研究过如:人流感病毒、柯萨奇病毒B3 (CVB3)、人多瘤病毒JCV等病毒。 siRNA在感染的早期阶段能有效地抑制病毒的复制,病毒感染能被针对病毒基因和相关宿主基因的siRNA所阻断,这些结果提示RNAi能胜任许多病毒防御与治疗。
、人多瘤病毒JCV等病毒。 siRNA在感染的早期阶段能有效地抑制病毒的复制,病毒感染能被针对病毒基因和相关宿主基因的siRNA所阻断,这些结果提示RNAi能胜任许多病毒防御与治疗。")
54
3.肿瘤治疗 用RNAi特异性地抑制癌基因、癌相关基因或突变基因的过度表达,使这类基因保持在静默或休眠状态,从而有望用这种新的手段治疗各种恶性肿瘤。 (1)RNAi可应用于敲除点突变激活的癌基因, (2)RNAi可应用于抑制插入基因及融合基因的表达, (3)RNAi可应用于抑制基因扩增。
RNAi可应用于抑制插入基因及融合基因的表达, (3)RNAi可应用于抑制基因扩增。")
55
4.药物开发 在药物开发过程中,用RNAi技术可以对靶基因的功能进行分析,有助于搞清药物的作用机制以及与基因编码产物,及与相应化合物的相互作用,从而更正确地发现药靶,开发更有效的候选药物。

56
总之, RNAi作为偶然发现的一种生物体的自然现象, 发现者本身可能并没有预料到其应用前景及潜在的价值, 但随着众多的研究人员的加入, 一个具有划时代意义的技术将有望使人类摆脱疾病的困扰.

57
siRNA实验技术 1. siRNA的设计 2. siRNA的制备 3. siRNA的转染

58
(一)siRNA的设计 (1)RNAi目标序列的选取原则
(1)从转录本(mRNA)的AUG起始密码开始,寻找“AA”二连序列,并记下其3'端的19个碱基序列,作为潜在的siRNA靶位点。 (2)将潜在的序列和相应的基因组数据库(人,或者小鼠,大鼠等等)进行比较,排除那些和其他编码序列同源的序列。 (3)选出合适的目标序列进行合成。通常一个基因需要设计多个靶序列的siRNA,以找到最有效的siRNA序列。
从转录本(mRNA)的AUG起始密码开始,寻找 AA 二连序列,并记下其3 端的19个碱基序列,作为潜在的siRNA靶位点。 (2)将潜在的序列和相应的基因组数据库(人,或者小鼠,大鼠等等)进行比较,排除那些和其他编码序列同源的序列。 (3)选出合适的目标序列进行合成。通常一个基因需要设计多个靶序列的siRNA,以找到最有效的siRNA序列。")
59
siRNA的设计 (2)阴性对照 一个完整的siRNA实验应该有阴性对照,作为阴性对照的siRNA应该和选中的siRNA序列有相同的组成,但是和mRNA没有明显的同源性。通常的做法是将选中的siRNA序列打乱,同样要检查结果以保证它和目的靶细胞中其他基因没有同源性。

60
(二)siRNA的制备 在非哺乳动物细胞的研究中,由于利用较长的双链就可以诱导RNAi而无须合成siRNA,因此设计RNAi实验的步骤比较简便。只要通过目的基因体外转录就可得到所需要的dsRNA,再通过浸泡、注射或转染靶细胞即可。然而在哺乳动物细胞内,较长的dsRNA(多于29个核苷酸)会导致非特异基因沉默,只有21个核苷酸大小的siRNA才能有效引发特异的基因沉默,所以RNAi的设计与非哺乳动物的有所不同。目前制备siRNA的方法主要有两类,即体外制备和体内表达,两者最大的不同在于:前者直接作用于RNA,而后者的对象则是DNA。根据具体工具不同可进一步划分为5种RNAi实验方法。
会导致非特异基因沉默,只有21个核苷酸大小的siRNA才能有效引发特异的基因沉默,所以RNAi的设计与非哺乳动物的有所不同。目前制备siRNA的方法主要有两类,即体外制备和体内表达,两者最大的不同在于:前者直接作用于RNA,而后者的对象则是DNA。根据具体工具不同可进一步划分为5种RNAi实验方法。")
61
siRNA的制备 化学合成法 体外转录法 siRNA表达载体 siRNA表达盒子 制备siRNA 的混合物

62
siRNA的制备 化学合成法 这种方法比任何其他方法都要昂贵,但是化学合成siRNA几乎不需要研究者的操作。
应遵循以下步骤:(1)扫描靶基因中的序列,记录每个3 ' 端19个核苷酸作为可能的siRNA靶位点;(2)筛选G/C含量较低的序列;(3)可能的序列要经适合的基因组数据库进行同源性分析,去除那些与其它基因明显同源的序列(推荐使用NCBI的BLAST);(4)最后得到长度为21个碱基的序列,从中选择2—4条进行测试。 除了实验成本过高以外,化学合成的另一个缺点就是周期较长。
扫描靶基因中的序列,记录每个3 端19个核苷酸作为可能的siRNA靶位点;(2)筛选G/C含量较低的序列;(3)可能的序列要经适合的基因组数据库进行同源性分析,去除那些与其它基因明显同源的序列(推荐使用NCBI的BLAST);(4)最后得到长度为21个碱基的序列,从中选择2—4条进行测试。 除了实验成本过高以外,化学合成的另一个缺点就是周期较长。")
63
siRNA的制备 体外转录法 体外转录法同样是体外制备siRNA,它要经济、快速许多,因为需要化学合成的是DNA。
体外转录合成的程序是,在设计好siRNA 序列后,先合成两条长度为29个碱基的DNA序列模板,其中包括5 ' 端与T7启动子引物3 ' 端互补的碱基(称引导序列),及下游编码选定的siRNA 的正义链或反义链的基因序列。然后将两条DNA模板与T7启动子引物退火结合,用DNA 聚合酶Klenow 合成dsDNA模板,再分别用T7 RNA聚合酶进行体外转录。最后将得到的产物经过单链专一的核酸酶消化,去除5 ' 末端的引导序列,随即得到需要的21个核苷酸的双链siRNA。经纯化后将其转入特定的载体中,转染进靶细胞就可以诱导目的基因沉默。另外也可用此法转录出长度为45—50个核苷酸的小发夹RNA ,它通过载体转染到细胞。shRNA 在细胞内会自动被加工成为siRNA,从而引发基因沉默。 利用此法得到的siRNA 产量有限,不适合制备大量siRNA以进行长期研究。
,及下游编码选定的siRNA 的正义链或反义链的基因序列。然后将两条DNA模板与T7启动子引物退火结合,用DNA 聚合酶Klenow 合成dsDNA模板,再分别用T7 RNA聚合酶进行体外转录。最后将得到的产物经过单链专一的核酸酶消化,去除5 末端的引导序列,随即得到需要的21个核苷酸的双链siRNA。经纯化后将其转入特定的载体中,转染进靶细胞就可以诱导目的基因沉默。另外也可用此法转录出长度为45—50个核苷酸的小发夹RNA ,它通过载体转染到细胞。shRNA 在细胞内会自动被加工成为siRNA,从而引发基因沉默。 利用此法得到的siRNA 产量有限,不适合制备大量siRNA以进行长期研究。")
64
siRNA的制备 制备siRNA 的混合物 该法克服了需要设计并测试许多siRNA序列的缺点。其原理是利用一种RNase 11酶消化dsRNA,用得到的混合物进行转染。 过程如下:(1)利用编码目的mRNA的长度为200—1000个碱基的DNA 作为转录模板,体外转录制备长的dsRNA;(2)在体外利用RNasem酶(或Dicer)消化长的dsRNA,产生siRNA的混合物;(3)纯化混合物,除去未消化的dsRNA。 存在诱发非特异性基因沉默的可能。
利用编码目的mRNA的长度为200—1000个碱基的DNA 作为转录模板,体外转录制备长的dsRNA;(2)在体外利用RNasem酶(或Dicer)消化长的dsRNA,产生siRNA的混合物;(3)纯化混合物,除去未消化的dsRNA。 存在诱发非特异性基因沉默的可能。")
65
siRNA的制备 siRNA表达载体 siRNA表达载体的特点是包含1种RNA聚合酶启动子(polm)以启动体内的转录,另外还有4—5个T的转录终止位点。常用RNA聚合酶启动子包括人和鼠的U6启动子和人H1启动子等3种。 操作过程为:首先订购两条编码目的小发卡siRNA 的寡聚DNA,经退火后克隆入所选载体的pol llI启动子下游。转入哺乳动物细胞,载体就能表达shRNA,随后被加工成siRNA,引发基因沉默。 由于涉及到克隆,且要对载体中包含嵌入物的区域进行测序,所以制备时间较长。而且质粒在某些不易转染的细胞中,转染效率较低。

66
siRNA的制备 siRNA表达盒子 siRNA表达盒子(SECs)是PCR引发的siRNA表达模板,与表达载体不同,它无需首先克隆入载体。
SECs依次包含RNA 聚合酶启动子、编码小发卡RNA 的模板DNA 序列和RNA 聚合酶终止位点。只要将插入EcoR I和HindllI限制性位点的RNA 聚合酶启动子和终止子的PCR引物,与合成好的模板DNA 序列在体外进行PCR扩增,即可得到大量SECs。将PCR扩增后的SECs插入表达载体中,直接转染至哺乳动物细胞,可以在体内引发功能性的siRNA 的表达,从而介导目的基因抑制。 SECs不像siRNA那样易于转染入细胞。另外,如果不克隆入质粒,SECs不易扩增。

67
siRNA的制备方法比较 制备时间 成本估计 序列测试 转染效果 最佳选择 化学合成 体外转录 混合物 表达载体 表达盒子 4天至2周
1至2天 1至2天 2至3周 1天 成本估计 最高 适中 低 适中 适中 序列测试 需要 需要 不需要 需要 需要 转染效果 好 好 好 一般 一般 最佳选择 确定最佳和高纯度序列 筛选测试序列 快速、低成本 长周期研究 筛选序列

68
(三)siRNA的转染 1.磷酸钙共沉淀 2.电穿孔法 3. DEAE-葡聚糖和polybrene 4.机械法 5.阳离子脂质体试剂
将制备好的siRNA,siRNA表达载体或表达框架转导至真核细胞中的方法主要有以下几种: 1.磷酸钙共沉淀 2.电穿孔法 3. DEAE-葡聚糖和polybrene 4.机械法 5.阳离子脂质体试剂

69
siRNA的转染 磷酸钙共沉淀 将氯化钙,RNA(或DNA)和磷酸缓冲液混合,沉淀形成包含DNA且极小的不溶的磷酸钙颗粒。磷酸钙-DNA复合物粘附到细胞膜并通过胞饮进入目的细胞的细胞质。沉淀物的大小和质量对于磷酸钙转染的成功至关重要。
和磷酸缓冲液混合,沉淀形成包含DNA且极小的不溶的磷酸钙颗粒。磷酸钙-DNA复合物粘附到细胞膜并通过胞饮进入目的细胞的细胞质。沉淀物的大小和质量对于磷酸钙转染的成功至关重要。 .")
70
siRNA的转染 电穿孔法 电穿孔通过将细胞暴露在短暂的高场强电脉冲中转导分子。将细胞悬浮液置于电场中会诱导沿细胞膜的电压差异,据认为这种电压差异会导致细胞膜暂时穿孔。电脉冲和场强的优化对于成功的转染非常重要,因为过高的场强和过长的电脉冲时间会不可逆地伤害细胞膜而裂解细胞。一般,成功的电穿孔过程都伴随高水平(50%或更高)的毒性。
的毒性。")
71
siRNA的转染 DEAE-葡聚糖和polybrene
带正电的DEAE-葡聚糖或polybrene多聚体复合物和带负电的DNA分子结合使得DNA可以结合在细胞表面。通过使用DMSO或甘油获得的渗透休克将DNA复合体导入。

72
siRNA的转染 机械法 转染技术也包括使用机械的方法,比如显微注射和基因枪(biolistic particle)。显微注射使用一根细针头将DNA,RNA或蛋白直接转入细胞质或细胞核。基因枪使用高压microprojectile将大分子导入细胞。
。显微注射使用一根细针头将DNA,RNA或蛋白直接转入细胞质或细胞核。基因枪使用高压microprojectile将大分子导入细胞。 .")
73
siRNA的转染 阳离子脂质体试剂 在优化条件下将阳离子脂质体试剂加入水中时,其可以形成微小的(平均大小约 nm)单层脂质体。这些脂质体带正电,可以靠静电作用结合到DNA的磷酸骨架上以及带负电的细胞膜表面。因此使用阳离子脂质体转染的原理与以前利用中性脂质体转染的原理不同。使用阳离子脂质体试剂,DNA并没有预先包埋在脂质体中,而是带负电的DNA自动结合到带正电的脂质体上,形成DNA-阳离子脂质体复合物。被俘获的DNA就会被导入培养的细胞。
单层脂质体。这些脂质体带正电,可以靠静电作用结合到DNA的磷酸骨架上以及带负电的细胞膜表面。因此使用阳离子脂质体转染的原理与以前利用中性脂质体转染的原理不同。使用阳离子脂质体试剂,DNA并没有预先包埋在脂质体中,而是带负电的DNA自动结合到带正电的脂质体上,形成DNA-阳离子脂质体复合物。被俘获的DNA就会被导入培养的细胞。")
74
siRNA的转染 1.纯化siRNA 2.避免RNA酶污染 3.健康的细胞培养物和严格的操作确保转染的重复性 4.避免使用抗生素
为了达到高的转染效率,在转染实验过程中,需要注意以下几点: 1.纯化siRNA 2.避免RNA酶污染 3.健康的细胞培养物和严格的操作确保转染的重复性 4.避免使用抗生素

75
siRNA实验技术 1. siRNA的设计 2. siRNA的制备 3. siRNA的转染

76
(一)siRNA的设计 (1)RNAi目标序列的选取原则
(1)从转录本(mRNA)的AUG起始密码开始,寻找“AA”二连序列,并记下其3'端的19个碱基序列,作为潜在的siRNA靶位点。 (2)将潜在的序列和相应的基因组数据库(人,或者小鼠,大鼠等等)进行比较,排除那些和其他编码序列同源的序列。 (3)选出合适的目标序列进行合成。通常一个基因需要设计多个靶序列的siRNA,以找到最有效的siRNA序列。
从转录本(mRNA)的AUG起始密码开始,寻找 AA 二连序列,并记下其3 端的19个碱基序列,作为潜在的siRNA靶位点。 (2)将潜在的序列和相应的基因组数据库(人,或者小鼠,大鼠等等)进行比较,排除那些和其他编码序列同源的序列。 (3)选出合适的目标序列进行合成。通常一个基因需要设计多个靶序列的siRNA,以找到最有效的siRNA序列。")
77
siRNA的设计 (2)阴性对照 一个完整的siRNA实验应该有阴性对照,作为阴性对照的siRNA应该和选中的siRNA序列有相同的组成,但是和mRNA没有明显的同源性。通常的做法是将选中的siRNA序列打乱,同样要检查结果以保证它和目的靶细胞中其他基因没有同源性。

78
(二)siRNA的制备 在非哺乳动物细胞的研究中,由于利用较长的双链就可以诱导RNAi而无须合成siRNA,因此设计RNAi实验的步骤比较简便。只要通过目的基因体外转录就可得到所需要的dsRNA,再通过浸泡、注射或转染靶细胞即可。然而在哺乳动物细胞内,较长的dsRNA(多于29个核苷酸)会导致非特异基因沉默,只有21个核苷酸大小的siRNA才能有效引发特异的基因沉默,所以RNAi的设计与非哺乳动物的有所不同。目前制备siRNA的方法主要有两类,即体外制备和体内表达,两者最大的不同在于:前者直接作用于RNA,而后者的对象则是DNA。根据具体工具不同可进一步划分为5种RNAi实验方法。
会导致非特异基因沉默,只有21个核苷酸大小的siRNA才能有效引发特异的基因沉默,所以RNAi的设计与非哺乳动物的有所不同。目前制备siRNA的方法主要有两类,即体外制备和体内表达,两者最大的不同在于:前者直接作用于RNA,而后者的对象则是DNA。根据具体工具不同可进一步划分为5种RNAi实验方法。")
79
siRNA的制备 化学合成法 体外转录法 siRNA表达载体 siRNA表达盒子 制备siRNA 的混合物

80
siRNA的制备 化学合成法 这种方法比任何其他方法都要昂贵,但是化学合成siRNA几乎不需要研究者的操作。
应遵循以下步骤:(1)扫描靶基因中的序列,记录每个3 ' 端19个核苷酸作为可能的siRNA靶位点;(2)筛选G/C含量较低的序列;(3)可能的序列要经适合的基因组数据库进行同源性分析,去除那些与其它基因明显同源的序列(推荐使用NCBI的BLAST);(4)最后得到长度为21个碱基的序列,从中选择2—4条进行测试。 除了实验成本过高以外,化学合成的另一个缺点就是周期较长。
扫描靶基因中的序列,记录每个3 端19个核苷酸作为可能的siRNA靶位点;(2)筛选G/C含量较低的序列;(3)可能的序列要经适合的基因组数据库进行同源性分析,去除那些与其它基因明显同源的序列(推荐使用NCBI的BLAST);(4)最后得到长度为21个碱基的序列,从中选择2—4条进行测试。 除了实验成本过高以外,化学合成的另一个缺点就是周期较长。")
81
siRNA的制备 体外转录法 体外转录法同样是体外制备siRNA,它要经济、快速许多,因为需要化学合成的是DNA。
体外转录合成的程序是,在设计好siRNA 序列后,先合成两条长度为29个碱基的DNA序列模板,其中包括5 ' 端与T7启动子引物3 ' 端互补的碱基(称引导序列),及下游编码选定的siRNA 的正义链或反义链的基因序列。然后将两条DNA模板与T7启动子引物退火结合,用DNA 聚合酶Klenow 合成dsDNA模板,再分别用T7 RNA聚合酶进行体外转录。最后将得到的产物经过单链专一的核酸酶消化,去除5 ' 末端的引导序列,随即得到需要的21个核苷酸的双链siRNA。经纯化后将其转入特定的载体中,转染进靶细胞就可以诱导目的基因沉默。另外也可用此法转录出长度为45—50个核苷酸的小发夹RNA ,它通过载体转染到细胞。shRNA 在细胞内会自动被加工成为siRNA,从而引发基因沉默。 利用此法得到的siRNA 产量有限,不适合制备大量siRNA以进行长期研究。
,及下游编码选定的siRNA 的正义链或反义链的基因序列。然后将两条DNA模板与T7启动子引物退火结合,用DNA 聚合酶Klenow 合成dsDNA模板,再分别用T7 RNA聚合酶进行体外转录。最后将得到的产物经过单链专一的核酸酶消化,去除5 末端的引导序列,随即得到需要的21个核苷酸的双链siRNA。经纯化后将其转入特定的载体中,转染进靶细胞就可以诱导目的基因沉默。另外也可用此法转录出长度为45—50个核苷酸的小发夹RNA ,它通过载体转染到细胞。shRNA 在细胞内会自动被加工成为siRNA,从而引发基因沉默。 利用此法得到的siRNA 产量有限,不适合制备大量siRNA以进行长期研究。")
82
siRNA的制备 制备siRNA 的混合物 该法克服了需要设计并测试许多siRNA序列的缺点。其原理是利用一种RNase 11酶消化dsRNA,用得到的混合物进行转染。 过程如下:(1)利用编码目的mRNA的长度为200—1000个碱基的DNA 作为转录模板,体外转录制备长的dsRNA;(2)在体外利用RNasem酶(或Dicer)消化长的dsRNA,产生siRNA的混合物;(3)纯化混合物,除去未消化的dsRNA。 存在诱发非特异性基因沉默的可能。
利用编码目的mRNA的长度为200—1000个碱基的DNA 作为转录模板,体外转录制备长的dsRNA;(2)在体外利用RNasem酶(或Dicer)消化长的dsRNA,产生siRNA的混合物;(3)纯化混合物,除去未消化的dsRNA。 存在诱发非特异性基因沉默的可能。")
83
siRNA的制备 siRNA表达载体 siRNA表达载体的特点是包含1种RNA聚合酶启动子(polm)以启动体内的转录,另外还有4—5个T的转录终止位点。常用RNA聚合酶启动子包括人和鼠的U6启动子和人H1启动子等3种。 操作过程为:首先订购两条编码目的小发卡siRNA 的寡聚DNA,经退火后克隆入所选载体的pol llI启动子下游。转入哺乳动物细胞,载体就能表达shRNA,随后被加工成siRNA,引发基因沉默。 由于涉及到克隆,且要对载体中包含嵌入物的区域进行测序,所以制备时间较长。而且质粒在某些不易转染的细胞中,转染效率较低。

84
siRNA的制备 siRNA表达盒子 siRNA表达盒子(SECs)是PCR引发的siRNA表达模板,与表达载体不同,它无需首先克隆入载体。
SECs依次包含RNA 聚合酶启动子、编码小发卡RNA 的模板DNA 序列和RNA 聚合酶终止位点。只要将插入EcoR I和HindllI限制性位点的RNA 聚合酶启动子和终止子的PCR引物,与合成好的模板DNA 序列在体外进行PCR扩增,即可得到大量SECs。将PCR扩增后的SECs插入表达载体中,直接转染至哺乳动物细胞,可以在体内引发功能性的siRNA 的表达,从而介导目的基因抑制。 SECs不像siRNA那样易于转染入细胞。另外,如果不克隆入质粒,SECs不易扩增。

85
siRNA的制备方法比较 制备时间 成本估计 序列测试 转染效果 最佳选择 化学合成 体外转录 混合物 表达载体 表达盒子 4天至2周
1至2天 1至2天 2至3周 1天 成本估计 最高 适中 低 适中 适中 序列测试 需要 需要 不需要 需要 需要 转染效果 好 好 好 一般 一般 最佳选择 确定最佳和高纯度序列 筛选测试序列 快速、低成本 长周期研究 筛选序列

86
(三)siRNA的转染 1.磷酸钙共沉淀 2.电穿孔法 3. DEAE-葡聚糖和polybrene 4.机械法 5.阳离子脂质体试剂
将制备好的siRNA,siRNA表达载体或表达框架转导至真核细胞中的方法主要有以下几种: 1.磷酸钙共沉淀 2.电穿孔法 3. DEAE-葡聚糖和polybrene 4.机械法 5.阳离子脂质体试剂

87
siRNA的转染 磷酸钙共沉淀 将氯化钙,RNA(或DNA)和磷酸缓冲液混合,沉淀形成包含DNA且极小的不溶的磷酸钙颗粒。磷酸钙-DNA复合物粘附到细胞膜并通过胞饮进入目的细胞的细胞质。沉淀物的大小和质量对于磷酸钙转染的成功至关重要。
和磷酸缓冲液混合,沉淀形成包含DNA且极小的不溶的磷酸钙颗粒。磷酸钙-DNA复合物粘附到细胞膜并通过胞饮进入目的细胞的细胞质。沉淀物的大小和质量对于磷酸钙转染的成功至关重要。 .")
88
siRNA的转染 电穿孔法 电穿孔通过将细胞暴露在短暂的高场强电脉冲中转导分子。将细胞悬浮液置于电场中会诱导沿细胞膜的电压差异,据认为这种电压差异会导致细胞膜暂时穿孔。电脉冲和场强的优化对于成功的转染非常重要,因为过高的场强和过长的电脉冲时间会不可逆地伤害细胞膜而裂解细胞。一般,成功的电穿孔过程都伴随高水平(50%或更高)的毒性。
的毒性。")
89
siRNA的转染 DEAE-葡聚糖和polybrene
带正电的DEAE-葡聚糖或polybrene多聚体复合物和带负电的DNA分子结合使得DNA可以结合在细胞表面。通过使用DMSO或甘油获得的渗透休克将DNA复合体导入。

90
siRNA的转染 机械法 转染技术也包括使用机械的方法,比如显微注射和基因枪(biolistic particle)。显微注射使用一根细针头将DNA,RNA或蛋白直接转入细胞质或细胞核。基因枪使用高压microprojectile将大分子导入细胞。
。显微注射使用一根细针头将DNA,RNA或蛋白直接转入细胞质或细胞核。基因枪使用高压microprojectile将大分子导入细胞。 .")
91
siRNA的转染 阳离子脂质体试剂 在优化条件下将阳离子脂质体试剂加入水中时,其可以形成微小的(平均大小约 nm)单层脂质体。这些脂质体带正电,可以靠静电作用结合到DNA的磷酸骨架上以及带负电的细胞膜表面。因此使用阳离子脂质体转染的原理与以前利用中性脂质体转染的原理不同。使用阳离子脂质体试剂,DNA并没有预先包埋在脂质体中,而是带负电的DNA自动结合到带正电的脂质体上,形成DNA-阳离子脂质体复合物。被俘获的DNA就会被导入培养的细胞。
单层脂质体。这些脂质体带正电,可以靠静电作用结合到DNA的磷酸骨架上以及带负电的细胞膜表面。因此使用阳离子脂质体转染的原理与以前利用中性脂质体转染的原理不同。使用阳离子脂质体试剂,DNA并没有预先包埋在脂质体中,而是带负电的DNA自动结合到带正电的脂质体上,形成DNA-阳离子脂质体复合物。被俘获的DNA就会被导入培养的细胞。")
92
siRNA的转染 1.纯化siRNA 2.避免RNA酶污染 3.健康的细胞培养物和严格的操作确保转染的重复性 4.避免使用抗生素
为了达到高的转染效率,在转染实验过程中,需要注意以下几点: 1.纯化siRNA 2.避免RNA酶污染 3.健康的细胞培养物和严格的操作确保转染的重复性 4.避免使用抗生素

93
回顾: PPT的制作: 讨论: 展示: 收获: 感谢:



,必须在活细胞内寄生并以复 制方式增殖的非细胞型微生物。 生物病毒微生物 原指一种动物来源的 毒素。 “virus” 一词源于 拉丁文。 病毒能增殖、遗传和演化,因而 具 有生命最基本的特征。>")
 ” 。 他的体内没有任何免疫系统, 没有任何抵御细菌、病毒的能力。>")
 体积微小,可以通过细菌滤器 ; 结构简单,只含一种类型的核酸 ; 专性细胞内寄生,以复制方式增殖的一类非细 胞型微生物。 对抗生素不敏感,对干扰素敏感。>")
染色体结构不同; ( 2 )原核生物具有正调控和负调控并重的特点,真核 生物目前已知的主要是正调控; ( 3 )原核生物的转录和翻译是相偶联的,真核生物的.>")















