Download presentation
Presentation is loading. Please wait.

1
第二节 噬菌体或病毒DNA Bacteriophage(phage)
是比细菌还小得多的微生物,噬菌体侵犯细菌,可以认为它是细菌里的“寄生虫”。 组成:蛋白质、核酸. 结构:蛋白质外壳、尾巴 尾巴上的微丝可以把噬菌体的DNA注入细菌内。 高效率的感染性能使外源基因高效导入受体细胞自主复制繁殖性能使外源基因在受体细胞中高效扩增 噬菌体:感染并裂解某种细菌的病毒

2
一、大肠杆菌的λ-噬菌体DNA (一) λ-噬菌体的生物学特性 1、由外壳包装蛋白和λ-DNA组成 2、 λ-DNA的物理图谱 48.5kb (1)线状双链DNA,两端各有一个12bp的互补单链(粘性末端,cohesive-end site),称λcos site ,粘性末端粘连接变成环状DNA。 λcos位点是噬菌体包装必需序列。
线状双链DNA,两端各有一个12bp的互补单链(粘性末端,cohesive-end site),称λcos site ,粘性末端粘连接变成环状DNA。 λcos位点是噬菌体包装必需序列。")
3
目前已经被确定的基因至少61种,一半为必需基因
(2)功能相近的基因在基因组中聚集在一起 目前已经被确定的基因至少61种,一半为必需基因 其中有一半左右参与了噬菌体生命周期的活动,我们称这类基因为λ噬菌体的必要基因;另一部分基因,当它们被外源基因取代之后,并不影响噬菌体的生命功能,我们称这类基因为λ噬菌体的非必要的基因。
功能相近的基因在基因组中聚集在一起. 目前已经被确定的基因至少61种,一半为必需基因. 其中有一半左右参与了噬菌体生命周期的活动,我们称这类基因为λ噬菌体的必要基因;另一部分基因,当它们被外源基因取代之后,并不影响噬菌体的生命功能,我们称这类基因为λ噬菌体的非必要的基因。")
5
λ噬菌体基因大致分为4个区: 结构区:A~ J19个基因,编码头、 尾部蛋白质
重组区: att(attachment)、int(intagrate) 及xis(excision) 调控区:启动子、终止子和N、CI基因 裂解区:O-R基因
、int(intagrate) 及xis(excision) 调控区:启动子、终止子和N、CI基因. 裂解区:O-R基因.")
6
3、感染周期(溶菌循环) λDNA复制早期:一个ori ,双向复制 晚期:滚环复制--多个λDNA分子形成线状多联体
吸附于大肠杆菌外膜上的LamB受体(正常功能是运转麦芽糖进入细胞内),麦芽糖可诱导这些受体的合成,故它也可促进λ噬菌体的吸附与感染(尾部吸附). (1)吸附 吸附于大肠杆菌外膜上的LamB受体(正常功能是运转麦芽糖进入细胞内),麦芽糖可诱导这些受体的合成,故它也可促进λ噬菌体的吸附与感染(尾部吸附)。 (2)DNA注入 (3)DNA复制 从单一ori区滚筒复制,形成线状多联体,λ-DNA上两个基因的产物激活复制的起始,但复制所需的蛋白因子系统则由宿主细胞提供。 (4)包装 包装蛋白在宿主细胞内合成,包装蛋白首先结合在cos区的附近,形成包装启动复合物,因此cos区对包装是至关重要的, 包装时由一个蛋白因子将cos区切开,从而保证只有一个DNA 线状负责被保证在一起,每个宿主细胞可包装多达100个成熟的λ-DNA分子,包装对DNA本身的性质无关,但对其大小要求比较严格,它只能包装λ-DNA分子的75%-105%,也就是说可以包装36.4kb-50.9kb范围内的DNA,包装好了的λ-DNA便形成成熟的噬菌体颗粒。 (5)裂解 每个细胞内容纳了多达100个成熟的噬菌体,这时两种负责裂解宿主细胞的蛋白被合成,细菌细胞被裂解,成熟的噬菌体释放出去,又去感染另外的宿主细胞,直至大肠杆菌细胞全部被裂解。 λDNA复制早期:一个ori ,双向复制 晚期:滚环复制--多个λDNA分子形成线状多联体
,麦芽糖可诱导这些受体的合成,故它也可促进λ噬菌体的吸附与感染(尾部吸附). (1)吸附 吸附于大肠杆菌外膜上的LamB受体(正常功能是运转麦芽糖进入细胞内),麦芽糖可诱导这些受体的合成,故它也可促进λ噬菌体的吸附与感染(尾部吸附)。 (2)DNA注入. (3)DNA复制 从单一ori区滚筒复制,形成线状多联体,λ-DNA上两个基因的产物激活复制的起始,但复制所需的蛋白因子系统则由宿主细胞提供。 (4)包装 包装蛋白在宿主细胞内合成,包装蛋白首先结合在cos区的附近,形成包装启动复合物,因此cos区对包装是至关重要的, 包装时由一个蛋白因子将cos区切开,从而保证只有一个DNA 线状负责被保证在一起,每个宿主细胞可包装多达100个成熟的λ-DNA分子,包装对DNA本身的性质无关,但对其大小要求比较严格,它只能包装λ-DNA分子的75%-105%,也就是说可以包装36.4kb-50.9kb范围内的DNA,包装好了的λ-DNA便形成成熟的噬菌体颗粒。 (5)裂解 每个细胞内容纳了多达100个成熟的噬菌体,这时两种负责裂解宿主细胞的蛋白被合成,细菌细胞被裂解,成熟的噬菌体释放出去,又去感染另外的宿主细胞,直至大肠杆菌细胞全部被裂解。 λDNA复制早期:一个ori ,双向复制. 晚期:滚环复制--多个λDNA分子形成线状多联体.")
7
包装 E 头部包装蛋白首先结合在cos区的附近,形成包装启动复合物,A蛋白切割cos位点。
包装 包装蛋白在宿主细胞内合成,包装蛋白首先结合在cos区的附近,形成包装启动复合物,因此cos区对包装是至关重要的, 包装时由一个蛋白因子将cos区切开,从而保证只有一个DNA 线状负责被保证在一起,每个宿主细胞可包装多达100个成熟的λ-DNA分子,包装对DNA本身的性质无关,但对其大小要求比较严格,它只能包装λ-DNA分子的75%-105%,也就是说可以包装36.4kb-50.9kb范围内的DNA,包装好了的λ-DNA便形成成熟的噬菌体颗粒。 (5)裂解 每个细胞内容纳了多达100个成熟的噬菌体,这时两种负责裂解宿主细胞的蛋白被合成,细菌细胞被裂解,成熟的噬菌体释放出去,又去感染另外的宿主细胞,直至大肠杆菌细胞全部被裂解。 头部包装蛋白首先结合在cos区的附近,形成包装启动复合物,A蛋白切割cos位点。
裂解 每个细胞内容纳了多达100个成熟的噬菌体,这时两种负责裂解宿主细胞的蛋白被合成,细菌细胞被裂解,成熟的噬菌体释放出去,又去感染另外的宿主细胞,直至大肠杆菌细胞全部被裂解。 头部包装蛋白首先结合在cos区的附近,形成包装启动复合物,A蛋白切割cos位点。")
8
4、溶源状态的建立 噬菌体感染大肠杆菌后,DNA直接整合到宿主细胞染色体DNA上,并不裂解细胞,这种情况称为溶源状态。
整合主要由cI和int基因的产物所激活,这两个基因的开放 与关闭取决于宿主细胞本身的性质。 cI基因:编码阻遏蛋白,是感染了λ噬菌体的寄主细胞进 入溶源化的必要条件。cI基因失活或缺失的λ噬菌体无法 使寄主细胞发生溶源化效应。 入侵E.coli后,可以按裂解或建立溶原两种生活方式生存和繁殖。侵犯细菌时,只是DNA侵入,然后利用细菌的酶来复制、转录、翻译。 DNA重组技术一般需要噬菌体处于溶源状态。

9
λ噬菌体DNA的整合与删除: 噬菌体基因组可以整合到大肠杆菌染色体DNA上,成为原噬菌体。
适当条件下,原噬菌体又可以脱离寄主染色体,重新变成独立的复制子,这种过程叫做原噬菌体的删除作用。 超感染免疫性 溶源性细菌,由于含有原噬菌体,故不能再 次被同种噬菌体感染。溶源性细菌所具有的这种 抗御同种噬菌体再感染的特性叫做超感染免疫性。 处于整合状态的噬菌体DNA称为原噬菌体

10
噬菌体的溶源和裂解

11
(二) λ-DNA载体的构建 1、缩短长度 野生型:上限51kb λ-DNA:48.5kb,外源基因2.5kb
λ-DNA上有40-50%的DNA是复制,裂解所不必需的,切除便提高装载量。 左侧区包括使噬菌体DNA成为一个成熟的、有外壳的病毒颗粒所需的全部基因,全长约20kb 右侧区内包含所有的调控因子、与DNA复制及裂解宿主菌有关的基因,这个区域约长12kb。 中间区域约长18kb,这一段DNA可以被外源置换而不会影响噬菌体λ裂解生长的能力。 左侧区包括使噬菌体DNA成为一个成熟的、有外壳的病毒颗粒所需的全部基因,全长约20kb 右侧区内包含所有的调控因子、与DNA复制及裂解宿主菌有关的基因,这个区域约长12kb。 中间区域约长18kb,这一段DNA可以被外源置换而不会影响噬菌体λ裂解生长的能力。置换型噬菌体λ是使用最广泛的载体。 根据切除的多少,将λ-DNA分为两大类载体: 插入型载体、取代型载体

13
插入型载体(insertion vector)
λgt10,可以携带8kb的DNA分子,插入位于cI基因内的单一的酶切位点EcoRI。插入失活导致裂解循环,从而很容易识别重组子。 λZAPII,含有多聚接头,可以使用六种不同的限制性内切酶插入约10kb的新的DNA分子,通过lacZ’基因的插入失活鉴定重组子,不能形成蓝色的噬菌斑。 必须携带标记基因 该类载体经改造后的长度为37kb,为包装的下限,它本身也能被包装,其允许的插入片段最大为13.9kb( kb)。由于该类载体重组与否均可包装,因而为区分重组子与非重组子必须携带标记基因。(0-13.9kb) λgt10,可以携带8kb的DNA分子,插入位于cI基因内的单一的酶切位点EcoRI。插入失活导致裂解循环,从而很容易识别重组子。 λZAPII,含有多聚接头,可以使用六种不同的限制性内切酶插入约10kb的新的DNA分子,通过lacZ’基因的插入失活鉴定重组子,不能形成蓝色的噬菌斑。 经改造后只具有一个可供外源DNA插入的克隆位点,长度为37kb,为包装的下限,它本身也能被包装,允许插入片段最大为14kb.
。由于该类载体重组与否均可包装,因而为区分重组子与非重组子必须携带标记基因。(0-13.9kb) λgt10,可以携带8kb的DNA分子,插入位于cI基因内的单一的酶切位点EcoRI。插入失活导致裂解循环,从而很容易识别重组子。 λZAPII,含有多聚接头,可以使用六种不同的限制性内切酶插入约10kb的新的DNA分子,通过lacZ’基因的插入失活鉴定重组子,不能形成蓝色的噬菌斑。 经改造后只具有一个可供外源DNA插入的克隆位点,长度为37kb,为包装的下限,它本身也能被包装,允许插入片段最大为14kb.")
14
取代型载体(substitution vector)
该类载体经改造后的长度约为40kb,但在非必需区域内含有两个相同的酶切口(如EcoRI、HindIII、或saλI),两者间的距离为14kb长,使用时用酶切开,分离去除这个14kb长的DNA片段,然后用外源DNA片段取代之。显而易见,这类载体的容量不仅比插入型载体大,而且有一个下限:40-14kb=26kb包装下限为36.4kb,因此其载装下限为10.4而上限为25.5kb。这类载体实际上已经不再需要标记基因,因为空载的载体DNA只有26kb,不可能被包装,因而无法进入受体细胞中去,当然这类载体的使用较为繁琐,现在商品化了的取代型载体已去除了中间片段 该类载体经改造后的长度约为40kb,但在非必需区域内含有两个相同的酶切口(如EcoRI、HindIII、或saλI),两者间的距离为14kb长,使用时用酶切开,分离去除这个14kb长的DNA片段,然后用外源DNA片段取代之。 具有成对的克隆位点,空载的载体DNA只26kb,不能被包装,无法进入受体细胞中去,不需要标记基因.
,两者间的距离为14kb长,使用时用酶切开,分离去除这个14kb长的DNA片段,然后用外源DNA片段取代之。显而易见,这类载体的容量不仅比插入型载体大,而且有一个下限:40-14kb=26kb包装下限为36.4kb,因此其载装下限为10.4而上限为25.5kb。这类载体实际上已经不再需要标记基因,因为空载的载体DNA只有26kb,不可能被包装,因而无法进入受体细胞中去,当然这类载体的使用较为繁琐,现在商品化了的取代型载体已去除了中间片段. 该类载体经改造后的长度约为40kb,但在非必需区域内含有两个相同的酶切口(如EcoRI、HindIII、或saλI),两者间的距离为14kb长,使用时用酶切开,分离去除这个14kb长的DNA片段,然后用外源DNA片段取代之。 具有成对的克隆位点,空载的载体DNA只26kb,不能被包装,无法进入受体细胞中去,不需要标记基因.")
15
1) Charon 系列 设计克隆外源 DNA 大片段的 取代型载体 2) EMBL 系列 设计克隆外源 DNA大片段 的 取代型载体。 优点:便于将外源DNA片段从重组体分子上切下。 3) 入-DASH 系列 含有 方向互为相反 的两套多 克隆位点接头序列, 优点: 便于多种 外源 DNA大片段的 取代重组。 4) 入-gt 系列 插入型表达 载体可用来克隆表达外源 c-DNA, 优点: 载体上温度敏感型阻遏物, 用来控制复制 及融合蛋白的表达。
 入-DASH 系列. 含有 方向互为相反 的两套多 克隆位点接头序列, 优点: 便于多种 外源 DNA大片段的 取代重组。 4) 入-gt 系列. 插入型表达 载体可用来克隆表达外源 c-DNA, 优点: 载体上温度敏感型阻遏物, 用来控制复制. 及融合蛋白的表达。")
16
应用: 插入型载体只能承受较小分子量(一般在10kb以内)的外源DNA片段的插入,广泛应用于cDNA及小片段DNA的克隆。 替换型载体可承受较大分子量的外源 DNA片段的插入,所以适用于克隆高等 真核生物的染色体DNA。

17
2、酶切位点的删除或增加 删除重复的酶切位点:野生型λDNA链上有 5个EcoRI位点和7个HindIII位点,不利于重 组操作,必须删除至1-2个. 为了便于各种来源的DNA片段的克隆,还 需要增加一些单一的酶切位点. 采用定点突变技术或甲基化酶处理必需区内的酶识别序列使其失活。

18
3、灭活某些与裂解周期有关的基因 将无义突变引入噬菌体裂解周期所需的基因, 如将头部包装蛋白基因的CAG突变成UAG。这些
噬菌体只能在具有特异校正基因编码产物的菌株内 繁殖。 当这种λDNA进入一般大肠杆菌菌株后,不能合 成有活性的头部蛋白,也就不能被包装和裂解细 菌,可阻止有害重组体的生物污染及扩散。 珀型突变(sup)是指由CAG(Gln)向UAG(stop)的突变。大肠杆菌的某些菌株含有特异性的校正基因,其编码产物校正tRNA能专一性地纠正这一突变。 基因工程实验中使用的受体是具有琥珀型突变体校正功能的菌株。
是指由CAG(Gln)向UAG(stop)的突变。大肠杆菌的某些菌株含有特异性的校正基因,其编码产物校正tRNA能专一性地纠正这一突变。 基因工程实验中使用的受体是具有琥珀型突变体校正功能的菌株。")
19
4、加装选择标记 选择标记主要有两类: 免疫功能类标记 颜色反应类标记 野生型λ-DNA上缺少合适的选择标记,因此加
与质粒不同,

20
(1)免疫功能失活标记 (cI筛选) 加装选择标记cI基因 cI基因编码一种阻止λ-噬菌体进入溶菌循环的阻遏物。
含有完整标记基因的λ-载体进入受体细胞后,建立溶源状态,细菌生长缓慢,形成混浊斑; 当外源DNA插入到标记基因中, 基因灭活, λ-重组分子便进入 溶菌循环,形成透明斑。 CⅠ基因的功能是促使噬菌体进入溶源化,但在正常情况下,它的噬菌斑是不清楚 的,有点浑浊。这是因为在λ噬菌体感染时,有少数细胞进入溶源化途径,这些细 胞生长在噬菌斑中,就造成了噬菌斑浑浊。由于cⅠ基因的产物——阻遏物的失活, 不会有溶源菌的形成,因此,形成的噬菌斑都是清亮的,很容易将重组体与非重组体区 别开来。cI 阻遏物的名字就是指在该基因中插入一段DNA 后会使噬菌斑的形态变得清 亮(c=clear)。
。")
21
(2)加装选择标记lacZ(蓝白斑筛选) lacZ基因编码β-半乳 糖苷酶,能催化无色的 X-gal生成蓝色化合物。
基因中,基因灭活,不能 合成蓝色化合物; 而空载体λ-DNA则产 生蓝色透明斑。

22
5、建立-噬菌体的体外包装系统 体外包装原理: λ噬菌体的头部和尾部的装配是分开进行的。
头部基因发生了突变的噬菌体只能形成尾部和头部蛋白所需的蛋白因子; 尾部基因发生了突变的噬菌体则只能形成头部和 尾部蛋白所需的蛋白因子。 λ-DNA重组分子需在体外人工包装成有感染力的噬菌体重组颗粒,方可高效导入受体细胞 用于体外包装的蛋白质可直接从感染了λ噬菌体的大肠杆菌中提 取,现已商品化。这些包装蛋白通常分为相互互补的两部分: 一部分缺少E组份,另一部分则缺少D组份。包装时,当且仅当 这两部分包装蛋白与重组λ-DNA分子混合后,包装才能有效进 行,任何一种蛋白包装液被重组λ-DNA污染后,均不能被包装 成有感染力的噬菌体颗粒,这也是基于安全而设计的 将这两种突变型的噬菌体的提取物混合起来,便能够在体外装配成有生物活性的噬菌体颗粒。

23
λDNA的体外包装作用,在离体条件下,将重组体DNA包入噬菌体等病毒颗粒中的过程,以形成有功能的病毒载体
不具有琥珀型突变体校正功能的菌株 λDNA的体外包装作用,在离体条件下,将重组体DNA包入噬菌体等病毒颗粒中的过程,以形成有功能的病毒载体。 sup-不具有琥珀型突变体校正功能的菌株 λDNA的体外包装作用,在离体条件下,将重组体DNA包入噬菌体等病毒颗粒中的过程,以形成有功能的病毒载体

24
可在体外包装成噬菌体颗粒,高效转染大肠杆菌 λ-DNA载体的装载能力为25 kb,远远大于质粒 的装载量 重组λ-DNA分子的筛选较为方便
λ-DNA载体适合克隆和扩增外源DNA片段,常用于 构建基因文库 在克隆载体基础上,为使插入的外源DNA片段有效转录翻译成多肽,装有强化外源基因表达的强启动子以及利于表达产物分泌、分离和纯化的元件,这种载体称为表达载体。 缺点: (1)需要包装比较麻烦,包装率又不稳定,购买包装蛋白的费用又高; (2)没有质粒的用途广泛。
需要包装比较麻烦,包装率又不稳定,购买包装蛋白的费用又高; (2)没有质粒的用途广泛。")
25
二、柯斯质粒(cosmid) (一)柯斯质粒的构建 考斯质粒:一类人工构建 的含有λDNA两端cos序 列和质粒复制子的特殊类 型的载体。
cos site-carrying plasmid”缩写而成的,其原意是指带有粘性末端位点(cos)的质粒。
的质粒。")
26
(二)柯斯质粒的特点 1、具有λ噬菌体的特性。 具有cos位点,能像 λ-DNA一样体外包装,并高效导入受体细胞。
2、具有质粒载体的持性。可以在受体细胞内 自由复制。 3、 具有高容量的克隆能力。可以装载比质粒或 λ-DNA大得多的外源DNA片段,40kb。 4、 便于筛选:携带质粒的选择标记 。 由于λ-DNA包装蛋白只识别粘性末端的一小段顺序cos区。将这段DNA与质粒连在一起,构 建的重组质粒,可装载外源DNA(45.5kb),它仍可象λ-DNA一样,在体外被包装成有感染 活性的噬菌体,进入细胞后,要通过质粒上的复制子进行复制。 柯斯质粒载体的分子仅具有一个复制起点,一两个选择记号和COS位点等三个组成部分,其分子量较小,一般只有5~7kb左右。因此,柯斯质粒载体的克隆极限可达45kb左右。 5、便于克隆:有质粒上的多种单一酶切位点。 6、不能体内包装,不裂解受体细胞
,它仍可象λ-DNA一样,在体外被包装成有感染. 活性的噬菌体,进入细胞后,要通过质粒上的复制子进行复制。 柯斯质粒载体的分子仅具有一个复制起点,一两个选择记号和COS位点等三个组成部分,其分子量较小,一般只有5~7kb左右。因此,柯斯质粒载体的克隆极限可达45kb左右。 5、便于克隆:有质粒上的多种单一酶切位点。 6、不能体内包装,不裂解受体细胞.")
27
利用Cos质粒进行克隆 “柯斯克隆” (cosmid cloning)
应用柯斯质粒载体,在大肠杆菌细胞中克隆大片段的真核基因组DNA技术,叫做柯斯克隆。 这种技术的理论依据是,在线性λ噬菌体DNΑ分子的每一端,都具有一段彼此互补的单链突出序列,即所谓的粘性末端(cos位点)。在λ噬菌体的正常生命周期中,会产生出由数百个λDNA拷贝组成的多连体分子。在此种分子中,前后两个λDNA基因组之间都是通过cos位点连接起来的。λ噬菌体具有的一种位点特异的切割体系(site-specific cutting system),叫做末端酶(terminase)或Ter体系,能识别两个相距适宜的cos位点,将多连体分子切割成λ单位长度的片段,并将它们包装到λ噬菌体头部中去。只有在被作用的λDNA分子具有两个cos位点,而且它们之间的距离保持在38~54kb的条件下,Ter体系才能对它们发生作用。 应用柯斯质粒作载体进行基因克隆的一般程序是:将外源DNA片段与柯斯质粒线性DNA分子进行体外连接反应。由此形成的连接产物群体中,有一定比例的分子是两端各有一个cos位点的长度为40kb左右的真核DNA片段,而且这两个cos位点在取向上是一样的,可作为λ噬菌体Ter功能的一种适用底物。当加入λ噬菌体的包装连接物时,它能把这些分子包装进λ噬菌体的头部,可以用来感染大肠杆菌(图5- “柯斯克隆” (cosmid cloning)
。在λ噬菌体的正常生命周期中,会产生出由数百个λDNA拷贝组成的多连体分子。在此种分子中,前后两个λDNA基因组之间都是通过cos位点连接起来的。λ噬菌体具有的一种位点特异的切割体系(site-specific cutting system),叫做末端酶(terminase)或Ter体系,能识别两个相距适宜的cos位点,将多连体分子切割成λ单位长度的片段,并将它们包装到λ噬菌体头部中去。只有在被作用的λDNA分子具有两个cos位点,而且它们之间的距离保持在38~54kb的条件下,Ter体系才能对它们发生作用。 应用柯斯质粒作载体进行基因克隆的一般程序是:将外源DNA片段与柯斯质粒线性DNA分子进行体外连接反应。由此形成的连接产物群体中,有一定比例的分子是两端各有一个cos位点的长度为40kb左右的真核DNA片段,而且这两个cos位点在取向上是一样的,可作为λ噬菌体Ter功能的一种适用底物。当加入λ噬菌体的包装连接物时,它能把这些分子包装进λ噬菌体的头部,可以用来感染大肠杆菌(图5- 柯斯克隆 (cosmid cloning)")
28
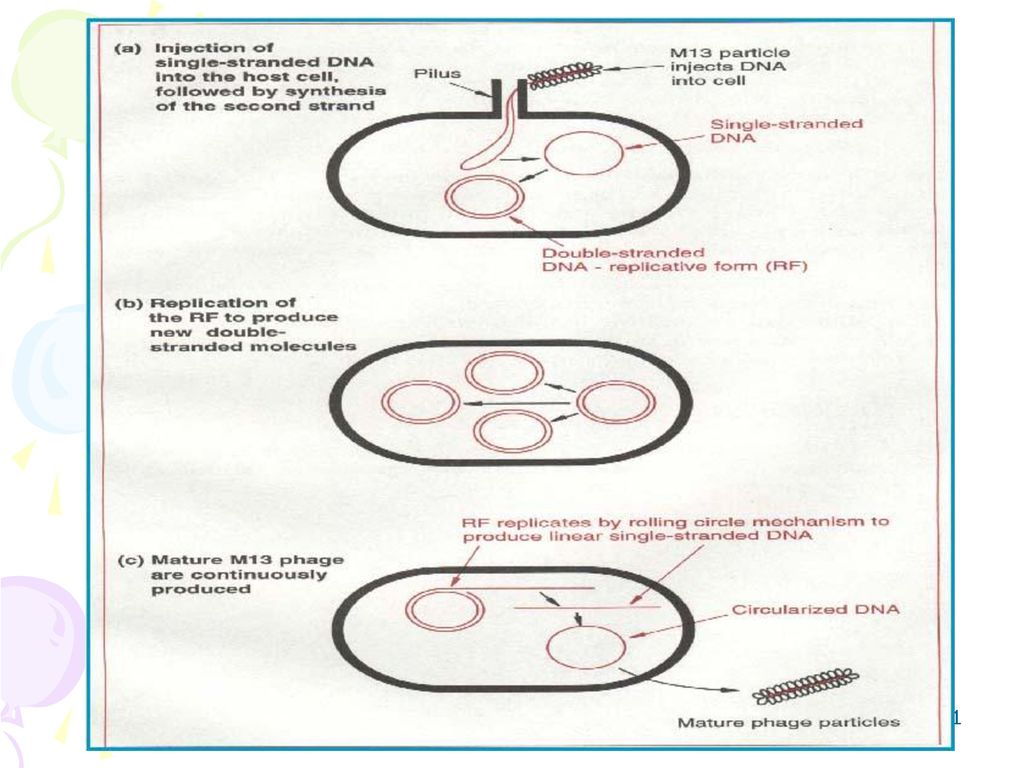
三、大肠杆菌的M13单链噬菌体DNA (一)生物结构 外形呈丝状由外壳包装蛋白和 正链DNA组成;不裂解宿主细 胞,但抑制其生长。
约有10个基因 M13、f1和fd,是一类丝状大肠杆菌噬菌体,它们都含有长度为6.4kb、彼此具有很高同源性的单链环状DNA分子。这些丝状噬菌体表现出一系列其它载体所不具备的优越性: 有时亦称为感染性的单链 M13噬菌体颗粒的外形呈丝状,在颗粒中包装的仅是(十)链的DNA,有时亦称为感染性的单链(图5-11)。 M13噬菌体首先吸附在F性须上,其吸附的位点看来是在性须的末端(图5-12)。噬菌体的基因组从性须末端进入到细胞的内部,进入细胞内部的M13(+)链DNA,便起到一种模板的作用,合成出互补的(一)链 DNA。由此形成的双链形式的M13DNA,称为复制型 DNA。它按θ形式进行几轮复制之后,基因 Ⅱ的产物便在RFDNA的正链待定位点上作切割反应,形成一个缺口。这样,M13基因组的扩增活动便正式开始启动。其基本特点是利用大肠杆菌的DNA聚合酶I,以环形的MI3(一)DNA为模板合成 M13(+)DNA。当DNA复制叉沿着模板DNA分子转移到复制终点时,在基因Ⅱ编码产物的作用下,新合成的(十)DNA便会被切除下去,并进一步环化形成单位长度的 M13基因组DNA(图5-13)。
链的DNA,有时亦称为感染性的单链(图5-11)。 M13噬菌体首先吸附在F性须上,其吸附的位点看来是在性须的末端(图5-12)。噬菌体的基因组从性须末端进入到细胞的内部,进入细胞内部的M13(+)链DNA,便起到一种模板的作用,合成出互补的(一)链 DNA。由此形成的双链形式的M13DNA,称为复制型 DNA。它按θ形式进行几轮复制之后,基因 Ⅱ的产物便在RFDNA的正链待定位点上作切割反应,形成一个缺口。这样,M13基因组的扩增活动便正式开始启动。其基本特点是利用大肠杆菌的DNA聚合酶I,以环形的MI3(一)DNA为模板合成 M13(+)DNA。当DNA复制叉沿着模板DNA分子转移到复制终点时,在基因Ⅱ编码产物的作用下,新合成的(十)DNA便会被切除下去,并进一步环化形成单位长度的 M13基因组DNA(图5-13)。")
29
M13噬菌体的基因组 编码3类蛋白质: ①复制蛋白(基因Ⅱ,Ⅴ和Ⅹ) ②形态发生蛋白(基因Ⅰ,Ⅳ和Ⅺ) ③结构蛋白(基因Ⅲ、Ⅵ、Ⅶ、Ⅷ和Ⅸ)
单链DNA,由6407碱基组成。 90%以上的序列可编码蛋白质,共有11个编码基因 基因之间的间隔区多为几个碱基。较大的间隔位于基因Ⅷ和基因Ⅲ以及基因Ⅱ和基因Ⅳ之间,其间有调节基因表达和DNA合成的元件。 编码3类蛋白质: ①复制蛋白(基因Ⅱ,Ⅴ和Ⅹ) ②形态发生蛋白(基因Ⅰ,Ⅳ和Ⅺ) ③结构蛋白(基因Ⅲ、Ⅵ、Ⅶ、Ⅷ和Ⅸ)
 ②形态发生蛋白(基因Ⅰ,Ⅳ和Ⅺ) ③结构蛋白(基因Ⅲ、Ⅵ、Ⅶ、Ⅷ和Ⅸ)")
30
(二)感染周期 θ复制 噬菌体的基因组从性须末端进入到细胞的内部,进入细胞内部的M13(+)链DNA,便起到一种模板的作用,合成出互补的(一)链 DNA。由此形成的双链形式的M13DNA,称为复制型 DNA。它按θ形式进行几轮复制之后,基因 Ⅱ的产物便在RFDNA的正链待定位点上作切割反应,形成一个缺口。这样,M13基因组的扩增活动便正式开始启动。其基本特点是利用大肠杆菌的DNA聚合酶I,以环形的MI3(一)DNA为模板合成 M13(+)DNA。当DNA复制叉沿着模板DNA分子转移到复制终点时,在基因Ⅱ编码产物的作用下,新合成的(十)DNA便会被切除下去,并进一步环化形成单位长度的 M13基因组DNA。
链DNA,便起到一种模板的作用,合成出互补的(一)链 DNA。由此形成的双链形式的M13DNA,称为复制型 DNA。它按θ形式进行几轮复制之后,基因 Ⅱ的产物便在RFDNA的正链待定位点上作切割反应,形成一个缺口。这样,M13基因组的扩增活动便正式开始启动。其基本特点是利用大肠杆菌的DNA聚合酶I,以环形的MI3(一)DNA为模板合成 M13(+)DNA。当DNA复制叉沿着模板DNA分子转移到复制终点时,在基因Ⅱ编码产物的作用下,新合成的(十)DNA便会被切除下去,并进一步环化形成单位长度的 M13基因组DNA。")
32
(三)M13 DNA载体的构建
M13 DNA载体的构建")
33
M13噬菌体作为载体具有几个重要的特点: ①M13噬菌体的感染与释放不会杀死宿主菌,仅导致宿主菌生长缓慢; ②M13噬菌体DNA在宿主菌内既可以是单链也可以是双链,通过感染或转化的方法能将M13噬菌体DNA导人宿主菌中; ③M13噬菌体的包装不受DNA大小的限制,其噬菌体颗粒的大小可随DNA的大小而改变,即使DNA的大小比本身DNA的大小超出6倍,仍能进行包装。

34
⑤M13噬菌体在用作载体时是利用其双链状态的RF DNA:单链DNA的酶切和连接是比较困难的。
⑥外源片段插入位点在基因Ⅱ和基因Ⅳ之间的508bp间隔区----M13不像λ噬菌体基因组那样含有较大的可替代区。它的基因组中绝大多数为必需基因,只有两个间隔区可用来插入外源DNA(基因Ⅱ/Ⅳ和基因Ⅷ/Ⅲ之间)。
。")
35
(四)M13DNA载体的特点 1、克隆的DNA片段以特定单链的形式输出受体细胞外,在DNA定向突变中非常有用. 2、M13重组体筛选简便
可以利用菌落的蓝白斑筛选转染的转化子 3、制备的单链DNA、双链RF-DNA M13噬菌体,不经体外包装,可以转染大肠杆菌受体。 通过感染或转化的方法能将M13DNA导人宿主菌中, 包装不受DNA大小的限制,其噬菌体颗粒的大小可随DNA的大小而改变. 包装不受DNA大小的限制,其噬菌体颗粒的大小可随DNA的大小而改变.即使DNA的大小比本身DNA的大小超出6倍,仍能进行包装。 4、缺点:包装能力有限,仅包装克隆DNA为1.5kb。

36
四、噬菌粒(phagemid or phasmid)
组成特点: 噬菌粒是一类人工构建的含有单链噬菌体包装序列、复制子以及质粒复制子、克隆位点、标记基因的特殊类型的载体。 像质粒那样在受体细胞中自主复制,克隆双链 DNA 能像M13 DNA那样体外包装,并高效转染 受体细胞,制备单链DNA 含有一个质粒的复制起点,因此在没有辅助噬菌体的情况下,克隆的外源基因可以像质粒一样按常规方法,复制形成大量的双链DNA分子; (viii)带有一个M13噬菌体的复制起点,所以在有辅助噬菌体感染的寄主细胞中,可以合成出单链DNA拷贝,并包装成噬菌体颗粒分泌到培养基中; (i)具有小分子量的共价、闭合、环形的基因组DNA,可克隆高达10kb的外源DNA片段,并易于进行体外分离与操作; (ii)编码有一个ampr基因作为选择记号,因此只有携带着pUCll8或pUCll9噬菌粒载体的大肠杆菌转化子细胞,才能够在含有氨千青霉素的培养基中生长,便于转化子的选择; (iii)拷贝数含量高,每个寄主细胞可高达500个,所以只要用少量的大肠杆菌细胞培养物,便可制备出大量的载体DNA; (iv)存在着一个多克隆位点区,因此许多种不同类型的外源DNA限制片段,不经修饰便可直接插入到载体分子上; (v)由于多克隆位点区阻断了大肠杆菌lacZ基因的5′-端编码区,故可按照Xgal-IPTG组织化学显色反应试验,筛选重组体分子; (vi)lacZ基因是置于lac启动子的控制之下,这样插入的外源基因(当其读码结构没有发生改变的情况下)便会以融合蛋白质形式表达,即产生出β半乳糖管酶与外源蛋白质的融合产物; 装载量比M13系列要大很多(10 kb) 重组操作简便,筛选容易
带有一个M13噬菌体的复制起点,所以在有辅助噬菌体感染的寄主细胞中,可以合成出单链DNA拷贝,并包装成噬菌体颗粒分泌到培养基中; (i)具有小分子量的共价、闭合、环形的基因组DNA,可克隆高达10kb的外源DNA片段,并易于进行体外分离与操作; (ii)编码有一个ampr基因作为选择记号,因此只有携带着pUCll8或pUCll9噬菌粒载体的大肠杆菌转化子细胞,才能够在含有氨千青霉素的培养基中生长,便于转化子的选择; (iii)拷贝数含量高,每个寄主细胞可高达500个,所以只要用少量的大肠杆菌细胞培养物,便可制备出大量的载体DNA; (iv)存在着一个多克隆位点区,因此许多种不同类型的外源DNA限制片段,不经修饰便可直接插入到载体分子上; (v)由于多克隆位点区阻断了大肠杆菌lacZ基因的5′-端编码区,故可按照Xgal-IPTG组织化学显色反应试验,筛选重组体分子; (vi)lacZ基因是置于lac启动子的控制之下,这样插入的外源基因(当其读码结构没有发生改变的情况下)便会以融合蛋白质形式表达,即产生出β半乳糖管酶与外源蛋白质的融合产物; 装载量比M13系列要大很多(10 kb) 重组操作简便,筛选容易.")
37
载体特点: 含有一个质粒的复制起点,因此在没有辅助噬菌体的情况下,克隆的外源基因可以像质粒一样按常规方法,复制形成大量的双链DNA分子;
具有小分子量的共价、闭合、环形的基因组DNA,可克隆高达10kb的外源DNA片段,并易于进行体外分离与操作; 编码有一个ampr基因作为选择记号,因此只有携带着pUCll8或pUCll9噬菌粒载体的大肠杆菌转化子细胞,才能够在含有氨千青霉素的培养基中生长,便于转化子的选择; 拷贝数含量高,每个寄主细胞可高达500个,所以只要用少量的大肠杆菌细胞培养物,便可制备出大量的载体DNA;

38
存在着一个多克隆位点区,因此许多种不同类型的外源DNA限制片段,不经修饰便可直接插入到载体分子上;
由于多克隆位点区阻断了大肠杆菌lacZ基因的5′-端编码区,故可按照Xgal-IPTG组织化学显色反应试验,筛选重组体分子; lacZ基因是置于lac启动子的控制之下,这样插入的外源基因(当其读码结构没有发生改变的情况下)便会以融合蛋白质形式表达,即产生出β半乳糖管酶与外源蛋白质的融合产物; 带有一个M13噬菌体的复制起点,所以在有辅助噬菌体感染的寄主细胞中,可以合成出单链DNA拷贝,并包装成噬菌体颗粒分泌到培养基中;
便会以融合蛋白质形式表达,即产生出β半乳糖管酶与外源蛋白质的融合产物; 带有一个M13噬菌体的复制起点,所以在有辅助噬菌体感染的寄主细胞中,可以合成出单链DNA拷贝,并包装成噬菌体颗粒分泌到培养基中;")
39
五、人工染色体克隆载体(artificial chromosome vector)
利用染色体的复制元件来驱动外源DNA片段复制的载体。 基本元件:含有质粒克隆载体所必需的第一受体(大肠杆菌)的质粒复制起始点; 第二受体(如酵母菌)染色体DNA着丝点、端粒和复制起始点的序列. 合适的选择标记基因。 人类、动物、植物的全基因组序列分析往往需要克隆数百 甚至上千kkbb 的DNA片段, 此时考斯质粒和噬菌粒载体的装载 量也远远不能满足需要。 将细菌接合因子或酵母菌染色体上的复制区、分配区、稳 定区与质粒组装在一起,即可构成染色体载体。当大片段的外 源DNA克隆在这些染色体载体上后,便形成重组人造染色体, 它能像天然染色体那样,在受体细胞中稳定的复制并遗传。 实际上是一种“穿梭”克隆载体, 在受体细胞中象质粒髙拷贝复制以便繁殖和保存 酵母人工染色体(YAC) 细菌人工染色体(BAC)
的质粒复制起始点; 第二受体(如酵母菌)染色体DNA着丝点、端粒和复制起始点的序列. 合适的选择标记基因。 人类、动物、植物的全基因组序列分析往往需要克隆数百. 甚至上千kkbb 的DNA片段, 此时考斯质粒和噬菌粒载体的装载. 量也远远不能满足需要。 将细菌接合因子或酵母菌染色体上的复制区、分配区、稳. 定区与质粒组装在一起,即可构成染色体载体。当大片段的外. 源DNA克隆在这些染色体载体上后,便形成重组人造染色体, 它能像天然染色体那样,在受体细胞中稳定的复制并遗传。 实际上是一种 穿梭 克隆载体, 在受体细胞中象质粒髙拷贝复制以便繁殖和保存. 酵母人工染色体(YAC) 细菌人工染色体(BAC)")
40
(一)酵母人工染色体(YAC) 装载量为350-400kb 含有的元件: 把酵母染色体与基因复制和表达有关的主要组件都组装
在质粒上,令质粒行使酵母chr的转录功能和复制功能。 含有的元件: 酵母4号染色体的复制起点和着丝粒序列 装载量为 kb 酵母系统的选择标记 a) YAC载体的结构和应用 pYAC3是在pBR322的基础上,插入了几个酵母基因。其中两个基因是URA3和TRP1,可以作为选择标记分别存在于YIp5, YRp7,就像在YRp7中一样,携带TRP1的同时也包含一个复制起点,但在pYAC3中这一片段扩展到CEN4,他是一段来自4号染色体着丝粒的DNA片段。TRP1-origin—CEN4片段已经包含了人工染色体组成三部分中的两部分。 第三部分端粒,有两段称为TEL的序列提供,并不是他们自身组成端粒序列,而是在引入酵母细胞核后,作为种子序列建立端粒。还有最后一部分没有提到的是SUP4,他在克隆实验中作为插入片段的选择标记基因。 利用pYAC3克隆的策略如下: YAC 载体的复制元件是其核心组成成分,其在酵母中复制的必需元件包括复制起点序列即自主复制序列(autonomously replicating sequence, ARS)、用于有丝分裂和减数分裂功能的着丝粒 (centromere , CEN)和两个端粒(TEL)。 YAC 人工染色体载体是利用酿酒酵母(Saccharomyces cerevisiae)的染色体的复制元件构建的载体,增加了普通大肠杆菌质粒载体的复制元件和选择标记,以便保存和增殖。 YAC 载体的复制元件是其核心组成成分,其在酵母中复制的必需元件包括复制起点序列即自主复制序列(autonomously replicating sequence, ARS)、用于有丝分裂和减数分裂功能的着丝粒 (centromere , CEN)和两个端粒(TEL) YAC 载体为能够满足自主复制、染色体在子代细胞间的分离及保持染色体稳定的需要,必须含有以下元件: ·端粒重复序列(telomeric repeat , TEL):定位于染色体末端一段序列,用于保护线状的 DNA 不被胞内的核酸酶降解,以形成稳定的结构。 ·着丝粒(centromere , CEN):有丝分裂过程中纺锤丝的结合位点, 使染色体在分裂过程中能正确分配到子细胞中 。在 YAC 中起到保证一个细胞内只有一个人工染色体的作用。如 pYAC4 使用的是酵母第四条染色体的着丝粒。 自主复制序列(autonomously replication sequences,ARS) 一段特殊的序列,含有酵母菌中 DNA 进行双向复制所必须的信号。 YAC 载体的选择标记主要采用营养缺陷型基因,如色氨酸、亮氨酸和组氨酸合成缺陷型基因 trp 1、 leu 2和 his 3、尿嘧啶合成缺陷型基因 ura3 等,以及赭石突变抑制基因 sup4 。 与 YAC 载体配套工作的宿主酵母菌(如AB1380)的胸腺嘧啶合成基因带有一个赭石突变 ade 2-1。带有这个突变的酵母菌在基本培养基上形成红色菌落,当带有赭石突变抑制基因 sup4 的载体存在于细胞中时,可抑制 ade 2-1基因的突变效应,形成正常的白色菌落。 有的可达 1Mb 以上,甚至达到 2Mb 质粒复制起点 质粒选择标记 四膜虫大核rDNA分子末端端粒重复序列
 YAC载体的结构和应用. pYAC3是在pBR322的基础上,插入了几个酵母基因。其中两个基因是URA3和TRP1,可以作为选择标记分别存在于YIp5, YRp7,就像在YRp7中一样,携带TRP1的同时也包含一个复制起点,但在pYAC3中这一片段扩展到CEN4,他是一段来自4号染色体着丝粒的DNA片段。TRP1-origin—CEN4片段已经包含了人工染色体组成三部分中的两部分。 第三部分端粒,有两段称为TEL的序列提供,并不是他们自身组成端粒序列,而是在引入酵母细胞核后,作为种子序列建立端粒。还有最后一部分没有提到的是SUP4,他在克隆实验中作为插入片段的选择标记基因。 利用pYAC3克隆的策略如下: YAC 载体的复制元件是其核心组成成分,其在酵母中复制的必需元件包括复制起点序列即自主复制序列(autonomously replicating sequence, ARS)、用于有丝分裂和减数分裂功能的着丝粒 (centromere , CEN)和两个端粒(TEL)。 YAC 人工染色体载体是利用酿酒酵母(Saccharomyces cerevisiae)的染色体的复制元件构建的载体,增加了普通大肠杆菌质粒载体的复制元件和选择标记,以便保存和增殖。 YAC 载体的复制元件是其核心组成成分,其在酵母中复制的必需元件包括复制起点序列即自主复制序列(autonomously replicating sequence, ARS)、用于有丝分裂和减数分裂功能的着丝粒 (centromere , CEN)和两个端粒(TEL) YAC 载体为能够满足自主复制、染色体在子代细胞间的分离及保持染色体稳定的需要,必须含有以下元件: ·端粒重复序列(telomeric repeat , TEL):定位于染色体末端一段序列,用于保护线状的 DNA 不被胞内的核酸酶降解,以形成稳定的结构。 ·着丝粒(centromere , CEN):有丝分裂过程中纺锤丝的结合位点, 使染色体在分裂过程中能正确分配到子细胞中 。在 YAC 中起到保证一个细胞内只有一个人工染色体的作用。如 pYAC4 使用的是酵母第四条染色体的着丝粒。 自主复制序列(autonomously replication sequences,ARS) 一段特殊的序列,含有酵母菌中 DNA 进行双向复制所必须的信号。 YAC 载体的选择标记主要采用营养缺陷型基因,如色氨酸、亮氨酸和组氨酸合成缺陷型基因 trp 1、 leu 2和 his 3、尿嘧啶合成缺陷型基因 ura3 等,以及赭石突变抑制基因 sup4 。 与 YAC 载体配套工作的宿主酵母菌(如AB1380)的胸腺嘧啶合成基因带有一个赭石突变 ade 2-1。带有这个突变的酵母菌在基本培养基上形成红色菌落,当带有赭石突变抑制基因 sup4 的载体存在于细胞中时,可抑制 ade 2-1基因的突变效应,形成正常的白色菌落。 有的可达 1Mb 以上,甚至达到 2Mb. 质粒复制起点. 质粒选择标记. 四膜虫大核rDNA分子末端端粒重复序列.")
41
克隆位点:位于sup4基因内 SuP4是一个赭石突变(UAA)抑制基因。 宿主酵母菌的胸腺嘧啶合成基因带有一个赭石突变 ade2。
有外源基因插入时,sup4基因遭破坏,则 形成红色菌落,十分容易挑选出有DNA插入片段的红色菌落,建成YAC文库。 酪氨酸 。带有这个突变的酵母菌在基本培养基上形成红色菌落,当带有赭石突变抑制基因 sup4 的载体存在于细胞中时,可抑制 ade 2-1基因的突变效应,形成正常的白色菌落。利用这一菌落颜色转变的现象,可用于筛选载体中含有外源 DNA 片段插入的重组子。 这是一种无意义突变。它是由于碱基代替换所产生的突变,这种突变使得原来对应于某种氨基酸的密码子变为赭石型终止密码子UAA,

42
酵母人工染色体的使用 载体首先被BamHI和SnaBI酶切将分子切成三块,BamHI片段被去掉,剩下两个臂,每一臂都已TEL作为末端,另一端是SnaBI位点。克隆的DNA必须是平端(SnaBI也是一个平端酶,识别序列是TACGTA)被连接在两个臂的中间就产生了人工染色体。利用原生质转化法将人工染色体引入酵母。受体酵母是利用一个双营养缺陷体trp1-ura3,载体上含有互补基因作为选择标记。转化后于基本培养基上培养,只有含有人工染色体的细胞可以正常生长。含有两个左臂或者两个右臂的染色体都不能正常生长,因为其中一个选择标记基因丢失,插入片段的鉴定可以通过SUP4基因的插入失活,白色的克隆是重组子,红色的不是。 对于 BamHⅠ切割后形成的微型酵母染色体,当用 EcoRⅠ或 SmaⅠ 切割抑制基因 sup4 内部的位点后形成染色体的两条臂,与外源大片段 DNA 在该切点相连就形成一个大型人工酵母染色体,通过转化进入到酵母菌后可象染色体一样复制,并随细胞分裂分配到子细胞中去,达到克隆大片段DNA 的目的。装载了外源 DNA 片段的重组子导致抑制基因 sup4 插入失活,从而形成红色菌落; 而载体自身连接后转入到酵母细胞后形成白色菌落。这些红色的装载了不同外源 DNA 片段的重组酵母菌菌落的群体就构成了 YAC 文库。 YAC文库装载的 DNA 片段的大小一般可达 kb,有的可达 1Mb 以上,甚至达到 2Mb Telomere
被连接在两个臂的中间就产生了人工染色体。利用原生质转化法将人工染色体引入酵母。受体酵母是利用一个双营养缺陷体trp1-ura3,载体上含有互补基因作为选择标记。转化后于基本培养基上培养,只有含有人工染色体的细胞可以正常生长。含有两个左臂或者两个右臂的染色体都不能正常生长,因为其中一个选择标记基因丢失,插入片段的鉴定可以通过SUP4基因的插入失活,白色的克隆是重组子,红色的不是。 对于 BamHⅠ切割后形成的微型酵母染色体,当用 EcoRⅠ或 SmaⅠ 切割抑制基因 sup4 内部的位点后形成染色体的两条臂,与外源大片段 DNA 在该切点相连就形成一个大型人工酵母染色体,通过转化进入到酵母菌后可象染色体一样复制,并随细胞分裂分配到子细胞中去,达到克隆大片段DNA 的目的。装载了外源 DNA 片段的重组子导致抑制基因 sup4 插入失活,从而形成红色菌落; 而载体自身连接后转入到酵母细胞后形成白色菌落。这些红色的装载了不同外源 DNA 片段的重组酵母菌菌落的群体就构成了 YAC 文库。 YAC文库装载的 DNA 片段的大小一般可达 kb,有的可达 1Mb 以上,甚至达到 2Mb. Telomere.")
43
3、YAC 染色体与宿主细胞的染色体大小相近,影响了YAC 载体的广泛应用
1、插入片段大,稳定性差 2、内部存在重组和嵌合现象 3、YAC 染色体与宿主细胞的染色体大小相近,影响了YAC 载体的广泛应用 YAC 载体功能强大,但有一些弊端。一个YAC中 克隆的DNA片段可能来自两个或多个不同的染色体上。)。插人的DNA较大,序列发 生重排,导致和原来染色体的序列不一致。重组很难检出 YAC结构和酵母天然染色体结构相 似,使用常规方法不易将YAC和酵母夭然染色体分开,要通过较长时间的PFE才能分离出YAC. 这主要表现在 3 个方面:首先,在 YAC 载体的插入片段会出现缺失 (deletion) 和基因重排 (rearrangement) 的现象。其次,容易形成嵌合体。嵌合就是在单个 YAC 中的插入片段由 2 个或多个的独立基因组片段连接组成。嵌合克隆约占总克隆的 5%~50% 。最后, YAC 染色体与宿主细胞的染色体大小相近,影响了 YAC 载体的广泛应用。 YAC 染色体一旦进入酿酒酵母细胞,由于其大小与内源的染色体的大小相近,就很难从中分离出来,不利于进一步分析
。插人的DNA较大,序列发. 生重排,导致和原来染色体的序列不一致。重组很难检出. YAC结构和酵母天然染色体结构相. 似,使用常规方法不易将YAC和酵母夭然染色体分开,要通过较长时间的PFE才能分离出YAC. 这主要表现在 3 个方面:首先,在 YAC 载体的插入片段会出现缺失 (deletion) 和基因重排 (rearrangement) 的现象。其次,容易形成嵌合体。嵌合就是在单个 YAC 中的插入片段由 2 个或多个的独立基因组片段连接组成。嵌合克隆约占总克隆的 5%~50% 。最后, YAC 染色体与宿主细胞的染色体大小相近,影响了 YAC 载体的广泛应用。 YAC 染色体一旦进入酿酒酵母细胞,由于其大小与内源的染色体的大小相近,就很难从中分离出来,不利于进一步分析.")
44
(二)细菌人工染色体(BAC) 在大肠杆菌性因子F质粒的基础上构建的, 在大肠杆菌受体菌维持单一拷贝,装载量范围 在50-300kb之间。
用于:克隆大型基因簇结构 构建动植物基因文库

45
Structure of a bacterial artificial chromosome (BAC), used for cloning large fragments of donor DNA. CMR is a selectable marker for chloramphenicol resistance. oriS, repE, parA, and parB are F genes for replication and regulation of copy number. cosN is the cos site from l phage. HindIII and BamHI are cloning sites at which foreign DNA is inserted. The two promoters are for transcribing the inserted fragment. The NotI sites are used for cutting out the inserted fragment.

46
(三)人工染色体克隆载体的应用 1、构建基因组文库 2、基因治疗 3、基因功能鉴定 b) YAC载体的使用
一些哺乳动物的基因大于100kb(如人类膀胱纤维基因有250kb),超过了大肠杆菌载体系统的承受范围,但正好在YAC载体的范围之内。最近的研究发现,在某些情况下,YACs可以在哺乳动物中表达,因此可以在基因存在的生物中研究基因的功能。 YACs在构建基因文库中非常重要,要知道,最高容量的大肠杆菌质粒可以插入300kb的片断,对于人的基因文库需要30000个克隆,而YACs可以克隆600kb的片段,一些特殊的类型可以携带1400kb片段,可以是人类基因文库的克隆是降至6500个。但不幸的是,这样的”mega-YACs”存在着不稳定性,克隆的DNA片段可以被重新排列。不管怎样,YACs在大规模的测序中例如人类基因组计划是非常有用的。 有的遗传病是多基因所致,称为连续多基因缺失综合症(Contiguous Gene Deletion Syndrome), MAC有能力容纳 整簇的基因,因而MAC能同时纠正多个位点不同基因缺失所致的遗传病。 由于高等真核生物的基因大多数是多外显子结构并且有长长的内含子,大型基因组片段可通过 YAC 载体转移到动物或动物细胞系中,进行功能研究。
,超过了大肠杆菌载体系统的承受范围,但正好在YAC载体的范围之内。最近的研究发现,在某些情况下,YACs可以在哺乳动物中表达,因此可以在基因存在的生物中研究基因的功能。 YACs在构建基因文库中非常重要,要知道,最高容量的大肠杆菌质粒可以插入300kb的片断,对于人的基因文库需要30000个克隆,而YACs可以克隆600kb的片段,一些特殊的类型可以携带1400kb片段,可以是人类基因文库的克隆是降至6500个。但不幸的是,这样的 mega-YACs 存在着不稳定性,克隆的DNA片段可以被重新排列。不管怎样,YACs在大规模的测序中例如人类基因组计划是非常有用的。 有的遗传病是多基因所致,称为连续多基因缺失综合症(Contiguous Gene Deletion Syndrome), MAC有能力容纳. 整簇的基因,因而MAC能同时纠正多个位点不同基因缺失所致的遗传病。 由于高等真核生物的基因大多数是多外显子结构并且有长长的内含子,大型基因组片段可通过 YAC 载体转移到动物或动物细胞系中,进行功能研究。")
47
练习题 1.列举质粒载体必须具备的基本特性。 2.某一质粒载体具有Tetr和Kanr的表型,在Kan抗性基因
内有一Bgl I的切点。现用 Bgl I切割该载体进行基因克 隆,问(1)涂皿时应加什么样的抗生素?(2)培养后长出的 菌落具有什么样的抗性基因型?(3)如何利用抗性变化筛选 到含有插入片段的重组体? 3.YAC载体具有什么样的功能性DNA序列?为什么在克隆大 片段时,YAC具有优越性? 4.如何将野生型的λ-噬菌体改造成为一个理想的载体? 5.什么是蓝白斑筛选法和c I 筛选法?
涂皿时应加什么样的抗生素 (2)培养后长出的. 菌落具有什么样的抗性基因型 (3)如何利用抗性变化筛选. 到含有插入片段的重组体 3.YAC载体具有什么样的功能性DNA序列 为什么在克隆大. 片段时,YAC具有优越性 4.如何将野生型的λ-噬菌体改造成为一个理想的载体 5.什么是蓝白斑筛选法和c I 筛选法")
Similar presentations


。>")
 一上一下二上二下三上 校訂必修校訂必修 英文 I 中文閱讀與寫作 I 計算機概論 I 體育 服務與學習教育 I 英文 II 中文閱讀與寫作 II 計算機概論 II 體育 服務與學習教育 II.>")







>")



DNA、RNA及蛋白质操作技术.>")





