Download presentation
Presentation is loading. Please wait.

1
养殖水域生态学 第七章 种群生态学

2
§1 种群的基本特征 种群的基本概念 种群的特征

3
一、种群的基本概念 1. 种群(Population): 种群就是占据某一地区一定空间中同种个体的集合。
种群过去译为人口、居群、繁群、族群、虫口等。种群的定义不同学者有着不同的理解,如: ①方宗熙(1975):“种群是由同一物种的若干个体组成的。种群是生活在同一地点,属于同一物种的一群个体。个体与种群的关系好比树木与森林的关系那样”。 ②钟章成(1987):“种群是占据某一地区的某个种的一群个体,其占据的边界往往与包括该种在内的生物群落界限相一致”。 ③迈尔(Mayr E,1970):“在现代分类学和种群遗传学的影响下,一个正在生物学中散布的用法,把种群一词限制在局部的种群,一个规定地区的具有可能交配的个体群”。
: 种群是由同一物种的若干个体组成的。种群是生活在同一地点,属于同一物种的一群个体。个体与种群的关系好比树木与森林的关系那样 。 ②钟章成(1987): 种群是占据某一地区的某个种的一群个体,其占据的边界往往与包括该种在内的生物群落界限相一致 。 ③迈尔(Mayr E,1970): 在现代分类学和种群遗传学的影响下,一个正在生物学中散布的用法,把种群一词限制在局部的种群,一个规定地区的具有可能交配的个体群 。")
4
④奥德姆(Odum EP,1971):“种群,指一群在同一地区同一物种的集合体,或者其他能交换遗传信息的个体集合体,具有许多特征,最好用统计函数来表示,但它描述的是集体特征而不是个体的特征。”。
这里要区分单一种群和混合种群、自然种群和实验种群,若生物属于同一种就是单一种群(single species population),若生物由数种混合在一起的就是混合种群(multi-species population);从生活的环境而言,在自然界的种群称为自然种群(natural population),实验室内的种群称为实验种群(laboratory population). 当从具体意义上用种群这个概念时,无论从空间上还是从时间上的界限,多少是随研究工作的方便而划分的。 总之,种群是同种生物在特定环境空间内的个体集群。在自然界,种群是生物种存在和进化的基本单位,是生物群落或生态系统的基本组成,同时也是生物资源开发、利用的对象。
,若生物由数种混合在一起的就是混合种群(multi-species population);从生活的环境而言,在自然界的种群称为自然种群(natural population),实验室内的种群称为实验种群(laboratory population). 当从具体意义上用种群这个概念时,无论从空间上还是从时间上的界限,多少是随研究工作的方便而划分的。 总之,种群是同种生物在特定环境空间内的个体集群。在自然界,种群是生物种存在和进化的基本单位,是生物群落或生态系统的基本组成,同时也是生物资源开发、利用的对象。")
5
二、种群的特征 种群是由个体组成的,但种群内的各个个体不是孤立的,而是通过种内关系组成的一个有机的统一体。种群决不等于个体的简单相加,从个体到种群是一个质的飞跃。个体有体积大小,而种群则是密度或数量多少;个体有生、死,而描述种群的生死过程则有出生率和死亡率,这是种群水平出现的新质。属于种群水平的新质还有年龄结构、性比等。个体与种群的关系是部分与整体的关系。自然种群有三大基本特征:

6
三大基本特征 1. 空间特征:指种群有一定的分布区域和型式。在分布范围内,环境条件适合种群的生存。分布中心条件最合适,边缘地区则波动较大。
2. 数量特征:种群密度随时间而变动。种群数量动态的形式多种多样,从理论上讲,大致可以分为如下几种形式:种群的增长;种群的动态平衡;种群的周期性波动;种群的不规则波动;种群数量的下降、种群的灭亡和种群灭绝。这些形式与各种群特有的出生率、死亡率、生长率和年龄结构等特性有密切关系。 3. 遗传特征:种群是一个基因库,有一定的遗传组成和遗传特征。种群在表观上的形态、生理、生态学现象,实质上是种群基因库在环境作用下适应、选择结果的反映。种群的遗传特征也使种群之间保持形态、生理和生态特征上的差异。

8
§2.种群的结构 种群结构的标志是种群密度,种群中个体的分布形式,年龄和性比和种群中个体的异质性。

9
一、种群密度 种群的核心问题是种群动态,种群动态就是研究种群密度在时间上和空间上的变动规律,种群的存在、增殖和发展都是以种群密度为特征的。密度是描述生物种群发展的重要参数。种群的群体特征之一是种群的密度(density),影响种群密度的四个参数是出生率(natality)、死亡率(mortality)、迁入率(immigration)和迁出率(emigration) 。
,影响种群密度的四个参数是出生率(natality)、死亡率(mortality)、迁入率(immigration)和迁出率(emigration) 。")
10
1、种群密度定义 种群密度是一个相对的度量,它是指单位面积或单位容积内有机体的量。这个量可以用个体数、生物量(湿重)、有机质干重或能量(KJ)等来表示。 以上标志各有作用,但每一个标志都不可能全面地反映种群密度的特点。如个体数便于表示种群内个体的相互距离;生物量可较客观地反映有机质浓度; 有机质干重和能量较好地反映了生物的质量。其中,个体数量和生物量是最常用的两项标志。由于密度是随时间推移而变化的,因此,密度所反映的通常是瞬时数量(instantaneous quantity)。 同一种群的密度用不同方法表示,结果可能大不相同。
。 同一种群的密度用不同方法表示,结果可能大不相同。")
11
2.阿利氏规律 种群密度过大会恶化本身的食物、氧气和其它生活条件。但是密度过稀也有不良影响。比如繁殖时难以找到异性和产生必需的性产物浓度等。因此每一物种都存在着最适的种群密度,并按照环境的具体条件而改变其最适密度,也就是说种群密度也是物种的适应特征之一,这就是所谓的阿利氏规律(Allee’s law),又称阿利氏原理(Allee’s principle)。即种群增长时存在两种情况,一种情况是当种群小时,存活率最高;另一种种群在中等大小时最有利,每一种群都有其自己的最适密度(optimum density),过疏和过密都会起到抑制作用。
,又称阿利氏原理(Allee’s principle)。即种群增长时存在两种情况,一种情况是当种群小时,存活率最高;另一种种群在中等大小时最有利,每一种群都有其自己的最适密度(optimum density),过疏和过密都会起到抑制作用。")
12
阿利氏原理

13
3.调节种群密度的因素 任何生物种群的数量都是有变化的。一般把数量大幅度的增减称为波动(fluctuation)小幅度的增减称为振动(oscillation)。对于自然种群,数量波动是最常见的。有的波动是规则的,称为周期性波动;有的波动是不规则的,称为非周期性波动。 种群数量变动反映着相互矛盾的两个过程——出生和死亡;迁入和迁出。

14
涉及种群调节的因素大致可区分为两大类:一类称非密度制约因素 (density – independent factor),其作用与种群本身的密度无关。另一类称密度制约 (density – dependent factor)因素,其作用是随着种群本身密度增加而增强。 通常食物、敌害、氧气、温度等是控制水生生物种群动态的主要因素。繁殖速度改变也常用以限制种群密度。 密度增大后常常发生个体的迁移与扩散。

15
二、种群中个体的空间分布类型 种群在一个区域的分布形式代表该种群在一定环境中的空间分布结构,同时又是种群的生物学特征对环境条件适应的结果。种群中个体的空间分布是关系到种群密度以及如何来判断密度量的大小问题。个体的空间分布一般可分为三类 :

16
1.均匀型(uniform)分布 适合于二项分布理论模型。种群中的个体之间作等距离分布。 这种情况是比较少见的。在肥水池塘中,当环境条件比较一致时,微型浮游生物的分布可能接近均匀型。
分布 适合于二项分布理论模型。种群中的个体之间作等距离分布。 这种情况是比较少见的。在肥水池塘中,当环境条件比较一致时,微型浮游生物的分布可能接近均匀型。")
17
2.随机型(random)分布 适合于Poisson 分布理论模型。个体的分布是偶然性的、无规则的。如森林地面的蜘蛛类,北欧的麋鹿。这种情况在水生生物中比较罕见,只是在环境特别一致的情况下,如在潮间带泥滩中的某些蛤类,常因海浪的冲刷而呈现随机分布。
分布 适合于Poisson 分布理论模型。个体的分布是偶然性的、无规则的。如森林地面的蜘蛛类,北欧的麋鹿。这种情况在水生生物中比较罕见,只是在环境特别一致的情况下,如在潮间带泥滩中的某些蛤类,常因海浪的冲刷而呈现随机分布。")
18
3.成群型(clumped)分布 在多数情形下,水生生物的空间分布并不随机,而是表现为不均匀的团状型。个体成群或团状型的形成或是由于生物的集群现象;或是由于环境的差异引起某一区域有利于生物的生活而另一些区域则不利;或是由于繁殖特性造成的,水生生物的聚集分布还常常由风和水流引起的。
分布 在多数情形下,水生生物的空间分布并不随机,而是表现为不均匀的团状型。个体成群或团状型的形成或是由于生物的集群现象;或是由于环境的差异引起某一区域有利于生物的生活而另一些区域则不利;或是由于繁殖特性造成的,水生生物的聚集分布还常常由风和水流引起的。")
19
以上这三种分布的划分并无绝对分明的界限,而成群分布是普遍而重要的。聚集分布在一定程度上强化了个体间在食物和空间上的竞争,但有利之处更多,表现出许多积极的生态学意义。
聚集分布型根据其分布特征又可进一步分为:核心分布型,适合于Neyman 分布理论模型;镶嵌分布型,适合于负二项分布理论模型;复合的分布型,如Polya-Eggenberger 核心分布等。 利用概率分布理论模型来确定种群的分布型式, 其优点是比较准确。上述每一种分布状态都有精确的参数来界定。

20
但是,频数分布理论仅反映种群在空间某一时刻相对静止的存在形式,却不能反映种群随时间变化所产生的聚集和扩散两种运动状态和信息。因此,一些学者从不同角度出发,提出了用不同聚集性指标来测验种群分布随时间变化的规律 。

21
Allee(1949)曾指出,动物的分布样式,会因为种内多度、密度的不同、以及理化和生物因素的影响而表现出很大差异。因此,在考虑种群分布形式时,分析环境因素对动物的作用机制是十分必要的。
曾指出,动物的分布样式,会因为种内多度、密度的不同、以及理化和生物因素的影响而表现出很大差异。因此,在考虑种群分布形式时,分析环境因素对动物的作用机制是十分必要的。")
22
三、种群的年龄分布和性比 (一)种群的年龄分布 (二)种群的性比
种群的年龄分布 (二)种群的性比")
23
(一)种群的年龄分布 种群的年龄结构是指种群中各年龄组(age class)在种群内所占的比例,即各年龄级的相对比率,称年龄比(age ratio)或年龄分布(age distribution)。 种群中的个体可分成三个生态时期(即繁殖前期、繁殖期和繁殖后期),这三期的比例是有变化的,因此,研究种群动态,不能离开年龄结构。 Lotaka(1925)从理论上证明,每一种类的种群倾向于形成一个稳定的年龄结构;如果这种稳定状态受到破坏,将在正常条件恢复时,回到原来的稳定状态。比较持久的改变,则将导致建立新的稳定分布,但被破坏程度大时不能恢复。 种群中的繁殖能力仅限于某些年龄,而死亡率在更大一些的年龄中机会更多些,从年龄组成中可以看出种群数量变化的动向。一般用年龄金字塔或年龄锥体(age pyramid)来表示种群的年龄结构。年龄锥体一般有三种类型:
,这三期的比例是有变化的,因此,研究种群动态,不能离开年龄结构。 Lotaka(1925)从理论上证明,每一种类的种群倾向于形成一个稳定的年龄结构;如果这种稳定状态受到破坏,将在正常条件恢复时,回到原来的稳定状态。比较持久的改变,则将导致建立新的稳定分布,但被破坏程度大时不能恢复。 种群中的繁殖能力仅限于某些年龄,而死亡率在更大一些的年龄中机会更多些,从年龄组成中可以看出种群数量变化的动向。一般用年龄金字塔或年龄锥体(age pyramid)来表示种群的年龄结构。年龄锥体一般有三种类型:")
24
种群年龄金字塔

25
1. 增长型种群:年龄锥体基部较宽,顶部狭窄,表示种群中有大量的幼体和极少数的老年个体,不仅出生率很高,大于死亡率,而且往往表现为指数增长,在这种情况下,种群后继世代的数量总是比前一世代多,使年龄结构图形表现出金字塔形,是一个正在迅速增长的种群。 2. 稳定型种群:年龄锥体基部和顶部的宽度相差不大。各年龄个体的百分比从幼龄到高龄逐渐减少。死亡率与出生率相近。即当种群的增长率逐渐下降,最终达到稳定的时候,繁殖前期个体与繁殖年龄的个体数量就会大体相等,而繁殖后期的个体数量仍维持较小的比例, 此即为稳定型的年龄结构的特点,其年龄结构呈钟形。 3. 衰退型种群:年龄锥体基部比较狭窄而顶部较宽,表示种群幼体比例很少,即繁殖前期的个体数量就会明显少于繁殖期和繁殖后期的个体数,老年个体比例较大。 此时的年龄结构就会表现为倒瓮形。

26
每一个“生态”年龄所占生命史的比例是随种类而异的。
水生动物的年龄组成,在很大程度上决定于各世代的成活率。 小型水生生物种群的年龄组成实际上决定于不同年龄个体被食害程度。

27
(二)种群的性比 种群的性别组成表现为雌雄个体的比例,它首先决定于种的特点。
种群性别组成和年龄组成一样,也是在环境条件变化的情形下,保证有效繁殖的一种适应。例如,当生活条件良好时,鱼类增加雌性个体以增强种群的繁殖力,反之则增加雄性个体以降低繁殖力。八目鳗种群中雌鱼的百分率即取决于种群的总量,种群总量越大,雌性比例越低(表6-2)。其它许多种鱼类也有类似现象。 还有一些鱼类(鲈鱼等)在饵料条件不佳时反而出现大量小型雌鱼,但这些雌鱼很早成熟,可供作大型雌鱼的食物,从而保证后者的顺利繁殖。
。其它许多种鱼类也有类似现象。 还有一些鱼类(鲈鱼等)在饵料条件不佳时反而出现大量小型雌鱼,但这些雌鱼很早成熟,可供作大型雌鱼的食物,从而保证后者的顺利繁殖。")
28
四、个体的异质性 种群中个体在形态和生理上的差别,称为个体的异质性。是物种最有效地利用生活资源的一种重要适应。如
兔子鱼同一种群的个体在鳃耙的密度和长度不同,以此扩大对饵料基础的利用效率。 年龄和性别的差异; 密度大时,同一卵袋孵化出的摇蚊幼虫大小参差不齐,分批变态,保证生活所需条件。 个体的异质性则扩大了整个种群的稳定性。环境恶化时,个体异质性增强。

29
§3.种群统计 种群数量是不断变化的,当内外环境有利时,种群数量增加;当条件不利时,种群数量减少,条件相对稳定时,种群变动的幅度也较小。种群数量取决于出生率和死亡率、迁入和迁出两组对立的过程,出生和迁入是使种群增加的因素;死亡和迁出则是种群减少的因素。

30
一、种群的度量 二、种群的出生率和死亡率 三、生命表、存活曲线和内禀增长率

31
一、种群的度量 不同种群的密度相差很大,如水中原生动物密度可达几亿个/m2;甲壳类可达几十万个/m2;而鱼类可能不足1尾。因此,不同种群的度量方法是不同的。严格地讲,密度(density)和数量(number)是有区别的。前者是指单位面积或体积内的某种生物种群的个体数,而后者强调的是某种生物种群的数量大小,实质上也隐含着面积或空间单位,否则,没有面积或空间单位的数量多少也就毫无意义。因此,在生态学中,种群的数量、大小(size)、密度是同义词,而多度或丰盛度(abundance)多指种群在量方面的特征。
和数量(number)是有区别的。前者是指单位面积或体积内的某种生物种群的个体数,而后者强调的是某种生物种群的数量大小,实质上也隐含着面积或空间单位,否则,没有面积或空间单位的数量多少也就毫无意义。因此,在生态学中,种群的数量、大小(size)、密度是同义词,而多度或丰盛度(abundance)多指种群在量方面的特征。")
32
(一)绝对密度的测定 1、总数量调查:计算某水体全部生活的某种生物的数量,此方法一般难度较大,一般不用。
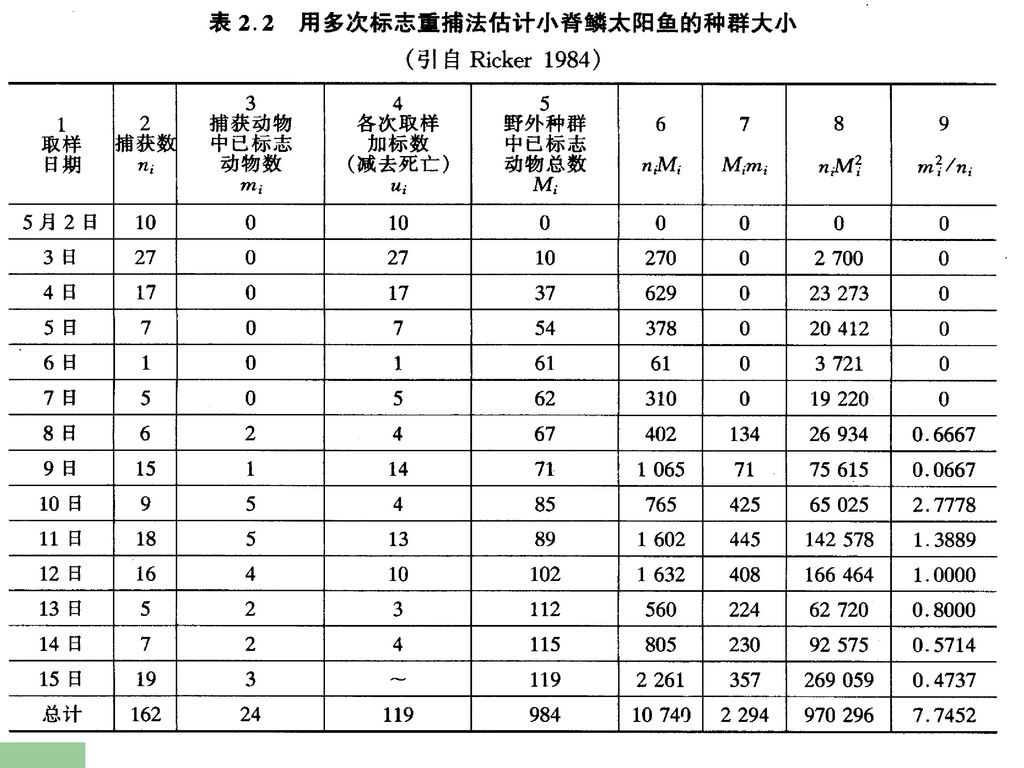
抽样调查法:在实际工作中一般多采用抽样调查,由样本来推断总体的规律性。因此,样本必须来自这个总体,而且必须具有代表性。 ①样方法(use of quadrat):通常计算若干样方(quadrat)或分样(subsample)中全部个体数。然后取其平均值来估计总体。 ②标志重捕法(mark-recapture method): 该法适用于统计鱼群数量。在水体中,捕捞一部分鱼将其标志,然后放回,经一定时间后进行重捕。根据重捕中标志数的比例,估计该水体中鱼的个体数。 ③去除取样法:用于调查小哺乳类。
:通常计算若干样方(quadrat)或分样(subsample)中全部个体数。然后取其平均值来估计总体。 ②标志重捕法(mark-recapture method): 该法适用于统计鱼群数量。在水体中,捕捞一部分鱼将其标志,然后放回,经一定时间后进行重捕。根据重捕中标志数的比例,估计该水体中鱼的个体数。 ③去除取样法:用于调查小哺乳类。")
33
去除取样法原理 是用相对统计法估计种群绝对量,其原理是:每次从生境中捕去已知数量的动物,因而影响下次捕捉。捕捉量的下降率与种群密度和已除去数量有关。本法必须满足下列条件: (1)捕捉活动不能影响未捕动物被捕的概率; (2)捕捉期间种群保持稳定,没有出生与死亡或迁入或迁出发生; (3)所有动物被捕率相等。
捕捉期间种群保持稳定,没有出生与死亡或迁入或迁出发生; (3)所有动物被捕率相等。")
35
去除取样法之一——回归分析法 以已捕量的累积量为横坐标X,依次捕捉量为纵坐标Y作图。该直线与横坐标的交点即为种群总数。或者以已捕量的累积量为自变量X,依次捕捉量为应变量Y,依最小二乘法得直线方程Y=a – bX。当Y=0时,X= a/b。 举例:如在某一种群中连续5次捕捉的诱捕量依次为65、43、34、18、12。以已捕获量的累积量为横坐标x,每次捕获量依次做纵坐标y作图(下图)目测绘出直线,该直线与横坐标的交点(x= 204)即为种群总数。或者以捕获量65、108、142、160、172为自变量x,依次捕获量65、43、34、18、12为应变量y,具最小二乘法得直线方程y = x,由此可知,当y=0时,x= 96.6/0.48=201,结果与目测法相近。
目测绘出直线,该直线与横坐标的交点(x= 204)即为种群总数。或者以捕获量65、108、142、160、172为自变量x,依次捕获量65、43、34、18、12为应变量y,具最小二乘法得直线方程y = x,由此可知,当y=0时,x= 96.6/0.48=201,结果与目测法相近。")
36
去除取样法——作图 方法1: 第1次捕捉 65 I 次捕捉数 43 34 18 12 204 已捕捉数累积量

37
去除取样法之二——三点法 三次捕捉t1,t2,t3,假定收集的数量与时间之间的指数关系可以从三次捕捉求出:
N = (n32-n1n2)/2n3-(n1+n2) 式中: n1、n2、n3各为t1,t2,t3时的累积捕捉量。如上例的累积捕捉量为65,108,142,160,172。如果t1=1, t2=5,则t3=(2+5)/2=3。代入上式得N=191。
/2n3-(n1+n2) 式中: n1、n2、n3各为t1,t2,t3时的累积捕捉量。如上例的累积捕捉量为65,108,142,160,172。如果t1=1, t2=5,则t3=(2+5)/2=3。代入上式得N=191。")
38
去除取样法之三——极大似然法

39
(二)相对密度的测定 相对密度即表示动物数量多少的相对指标。一般在无法或难以确定种群的绝对密度时采用。一般可用:
(1)多 或 ++++或abundunt(A.) (2)一般或 +++ 或common ( C.) (3)少 或 ++ 或 occasionic(O.) (4)极少或 + 或 rare ( R.)
多 或 ++++或abundunt(A.) (2)一般或 +++ 或common ( C.) (3)少 或 ++ 或 occasionic(O.) (4)极少或 + 或 rare ( R.)")
40
二、种群的出生率和死亡率 率(rate)是在生态学定量研究中的一个重要概念,所谓率是两组事物之间的数值比例。在生态学中,一般是以时间单位来表示率,即以变化量除以时间。如某种动物100个个体,一年中死亡20个,则该动物年死亡率为20%。
是在生态学定量研究中的一个重要概念,所谓率是两组事物之间的数值比例。在生态学中,一般是以时间单位来表示率,即以变化量除以时间。如某种动物100个个体,一年中死亡20个,则该动物年死亡率为20%。")
41
(一)出生率(natality) 出生率泛指任何生物产生新个体的能力。不论这些新个体是通过分裂、孵化、再生等那一种方法,都可用出生率这一术语加以概括。 出生率指种群中产生新个体的速率,可以用绝对值和相对值表示。种群出生率常常分为最大出生率(maximum natality)或潜在出生率(potential natality)和实际出生率(realized natality)或生态出生率(ecological natality)。最大出生率是指种群处于最适条件下(不受任何生态因子的限制)的出生率;实际出生率则指在特定环境条件下的种群出生率。 需要说明的是,这里指的出生率都是对种群而言,即种群的平均繁殖能力。至于种群中某些个体出现的超常生殖能力则不能代表种群的最大出生率。
或潜在出生率(potential natality)和实际出生率(realized natality)或生态出生率(ecological natality)。最大出生率是指种群处于最适条件下(不受任何生态因子的限制)的出生率;实际出生率则指在特定环境条件下的种群出生率。 需要说明的是,这里指的出生率都是对种群而言,即种群的平均繁殖能力。至于种群中某些个体出现的超常生殖能力则不能代表种群的最大出生率。")
42
1.影响种群出生率高低的因素 种群出生率的高低决定于繁殖方式、生殖量、生殖率和发育速度。

43
(1) 繁殖方式 水生生物的繁殖方式是多样的,不但不同种类有不同的繁殖方式,同一种类也可能有几种繁殖方式。概况起来,有无性繁殖、有性繁殖和两种方式交替进行三种类型。后一种情况是有机体最有效地利用生存条件的一种生态生理适应。例如,硅藻以无性繁殖方式迅速增殖,但经历多次繁殖后个体小型化,这时,可通过有性生殖来复大。许多水生动物在条件不利时进行有性生殖,形成遗传性较多样化的后代,以增强种群的适应能力;而在条件好转时进行无性生殖或单性生殖,其后代的遗传性较单纯,但繁殖速度加快。按照生活条件,单性世代和两性世代的数目随食物和温度的变化而变化,相应地在水生生物中可划分为单周期种(一年有一个有性世代)、双周期种(一年有两个有性世代)、多周期种(一年有多个有性世代)和无周期种(几乎全为单性生殖)。
、双周期种(一年有两个有性世代)、多周期种(一年有多个有性世代)和无周期种(几乎全为单性生殖)。")
44
(2)生殖量 生殖量指每次生殖时产出后代的数量,可以用绝对生殖量-每个雌体每次产卵数(产幼数)和相对生殖量-雌性的单位体重每次产卵数来表示。
不同种类水生动物的生殖量差别很大。 大多数动物的生殖量随年龄或个体大小而增高,以后又随个体开始衰老而下降。 食物的质量和丰度对生殖量有重要的影响。 环境的非生物因素也影响着生殖量。 鱼类和其它水生动物可以通过减少卵黄储备或使卵黄浓缩而缩小卵的形状,从而增加怀卵数。 动物一生的产卵(产幼)总数称为总生殖量。总生殖量决定于生殖量和一生中的生殖次数。
总数称为总生殖量。总生殖量决定于生殖量和一生中的生殖次数。")
45
(3)发育速度 发育的快慢决定着性成熟的早晚和单位时间内的生殖次数。 温度是决定发育速度和生殖率(动物单位时间内生殖次数)的主要因子。
发育速度 发育的快慢决定着性成熟的早晚和单位时间内的生殖次数。 温度是决定发育速度和生殖率(动物单位时间内生殖次数)的主要因子。")
46
2.出生率的计算 出生率还可分为绝对出生率(B,absolute birth rate)和相对出生率(b, relative natality)。其计算公式为: B = △Nn/△t b = △Nn/N△t 式中,N为种群的大小,△Nn种群内新产生的个体数,△t为时间改变量或单位时间。 在水生无脊椎动物的种群生态的研究中,更常用的是期限出生率(finite birth rate)和瞬时出生率(instantaneous birth rate),通常也以B和b表示。前者是指某一时间间隔中的平均出生率;后者则是指某一瞬间的出生率。两者的计算式为: B=E/D b = ln(1+B)或=ln(1+E) /D 式中:E为卵数与雌体数的比率(卵数/雌体数);D 为卵的发育时间(d)。 如果瞬时出生率(b)与瞬时死亡率(d)相等,则b=B;如果d=0,则b=ln(1+B);假如b≠d>0,则b=ln(1+E)/D。
和瞬时出生率(instantaneous birth rate),通常也以B和b表示。前者是指某一时间间隔中的平均出生率;后者则是指某一瞬间的出生率。两者的计算式为: B=E/D. b = ln(1+B)或=ln(1+E) /D. 式中:E为卵数与雌体数的比率(卵数/雌体数);D 为卵的发育时间(d)。 如果瞬时出生率(b)与瞬时死亡率(d)相等,则b=B;如果d=0,则b=ln(1+B);假如b≠d>0,则b=ln(1+E)/D。")
48
(二)死亡率和存活率 死亡率和出生率相反,表示因死亡而离开种群的个体数, 也可以用绝对值和相对值来表示。绝对死亡率指种群在单位时间内死亡的总数;相对死亡率指死亡个体数对原有种群数量的百分比。未死亡留在种群中的个体数与原有种群总数量的百分比称存活率。 种群的死亡率也可以分为最低死亡率(minimium mortality)和实际死亡率(生态寿命)。最低死亡率指种群在最适环境条件下,种群中的个体都是由于老年而死亡,即动物都活到生理寿命(phisiological longevity)才死亡。种群的生理寿命是指种群处于最适条件下的平均寿命,而不是某个特殊个体可能具有的最长寿命。实际死亡率则是指种群在特定环境条件下的平均死亡率,亦即种群活到生态寿命时的死亡率。生态寿命是指种群在特定环境条件下的实际平均寿命。换言之,只有一部分个体才能活到生理寿命,多数则死于捕食者、疾病、不良气候等原因。
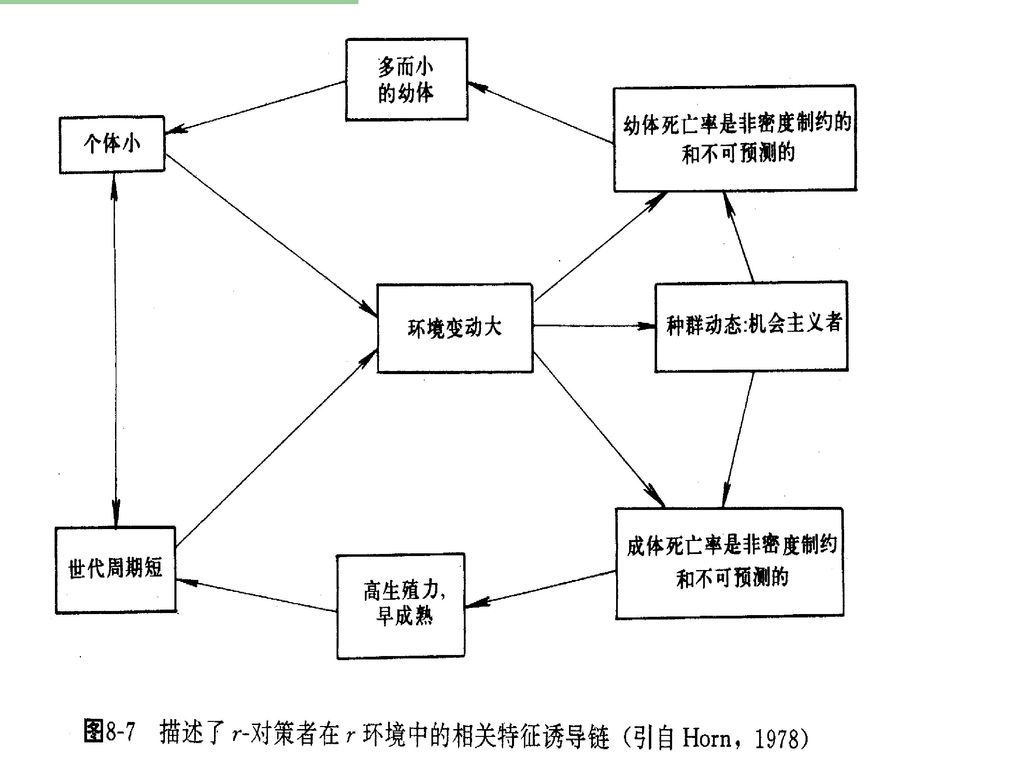
和实际死亡率(生态寿命)。最低死亡率指种群在最适环境条件下,种群中的个体都是由于老年而死亡,即动物都活到生理寿命(phisiological longevity)才死亡。种群的生理寿命是指种群处于最适条件下的平均寿命,而不是某个特殊个体可能具有的最长寿命。实际死亡率则是指种群在特定环境条件下的平均死亡率,亦即种群活到生态寿命时的死亡率。生态寿命是指种群在特定环境条件下的实际平均寿命。换言之,只有一部分个体才能活到生理寿命,多数则死于捕食者、疾病、不良气候等原因。")
49
在实验培养条件下,各种水生生物的最大寿命虽然相差很大,但是较之它们在自然界中可活的时间都大得多。这是因为,在天然水体中,水生生物很少因本身的衰老而死亡,绝大多数是由于被捕食、寄生或不利的理化条件影响,在不同的发育阶段死亡而离开种群的。 在自然条件下调查野生动物的种群死亡率比较困难,一般通过存活率而间接获得。

50
1、生物对减少死亡率的适应 ⑴胚胎发育期对降低死亡率的适应 ①创造有利的呼吸和温度条件: A.把卵产在富于氧气的场所。
B.把卵产在母体上孵化,此时,随着动物运动,卵也经常受新鲜水流的冲洗。从而有利呼吸。 C.胚胎发育期在较好的温度条件下进行。 ②防御敌害免被取食 软体动物和昆虫的卵块、甲壳类、鱼类的卵都透明无色,不被敌害发现;卵膜可保护卵免受微生物侵蚀;亲体主动守护卵等。 ③休眠卵:轮虫、枝角类、卤虫等。

51
生物对减少死亡率的适应-2 ⑵生长特性的适应 生物的生长是其生命过程中体质的积累,表现为体积的增大和体重的增加,是减少死亡率的一种适应。
水生生物的生长具有连续性和周期性,连续性生长是恒温动物和生活条件稳定的变温动物的特性,温带及高纬度水体的变温动物每年生长期有长有短。 ⑶构造和行为的适应 外覆物:贝壳、几丁质或角质外皮,棘刺等; 毒 素:有毒鱼卵,小球藻素,鱼毒素; 保护色:鱼类,摇蚊幼虫 躲避处:水草、岩石、土壤等; 活动性:快速运动。

52
2.死亡率的计算 死亡率也可分为绝对死亡率(D)和相对死亡率(d),计算公式分别为: D = △Nn/△t d =△Nn/N△t
在水生无脊椎动物的种群生态的研究中也经常用期限死亡率(finite death rate)和瞬时死亡率(instantaneous death rate),也分别以 D和d表示。前者是指某一时间间隔中的平均死亡率;后者则是指某一瞬间的死亡率率。两者的计算式为: D = 1 – e-d -d = ln(1 - D)
和瞬时死亡率(instantaneous death rate),也分别以 D和d表示。前者是指某一时间间隔中的平均死亡率;后者则是指某一瞬间的死亡率率。两者的计算式为: D = 1 – e-d. -d = ln(1 - D)")
53
三、生命表、存活曲线和内禀增长率 (一)生命表 (二)存活曲线和期望寿命 (三)特定年龄生育率(mx)和净生殖率(R0)
生命表 (二)存活曲线和期望寿命 (三)特定年龄生育率(mx)和净生殖率(R0)")
54
(一)生命表 生命表(life table)是描述种群死亡过程的一览表。生命表应用较广,作为生态学工作者应该学会编制生命表的具体方法。
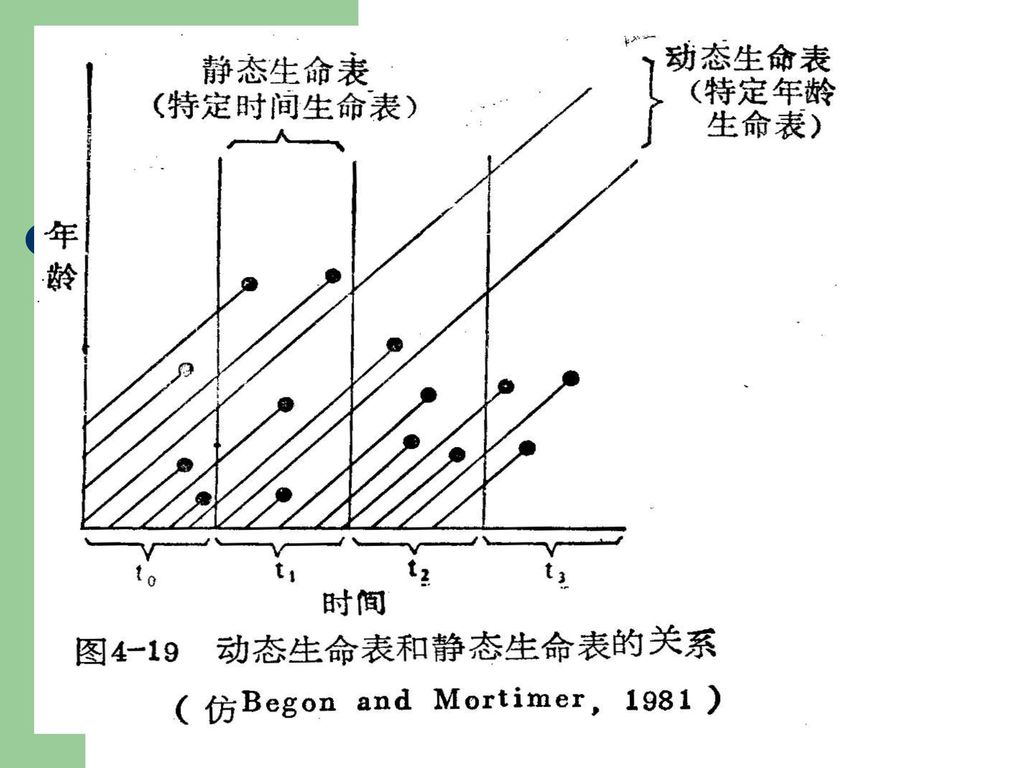
生命表数据的搜集有两种: 1.静态生命表:根据某一特定时间,对种群作一年龄分布的调查,根据其结果而编制。这一类生命表的各年龄组都是在不同的年中经历过来的。当然假定种群的年龄组成不变,且年复一年种群经历的环境不变。在鱼类和鸟类研究中常用。 2. 动态生命表:即同时观察一群同时出生的生物存活的数据来编制生命表。如藤壶的生命表。枝角类的生命表亦然。

56
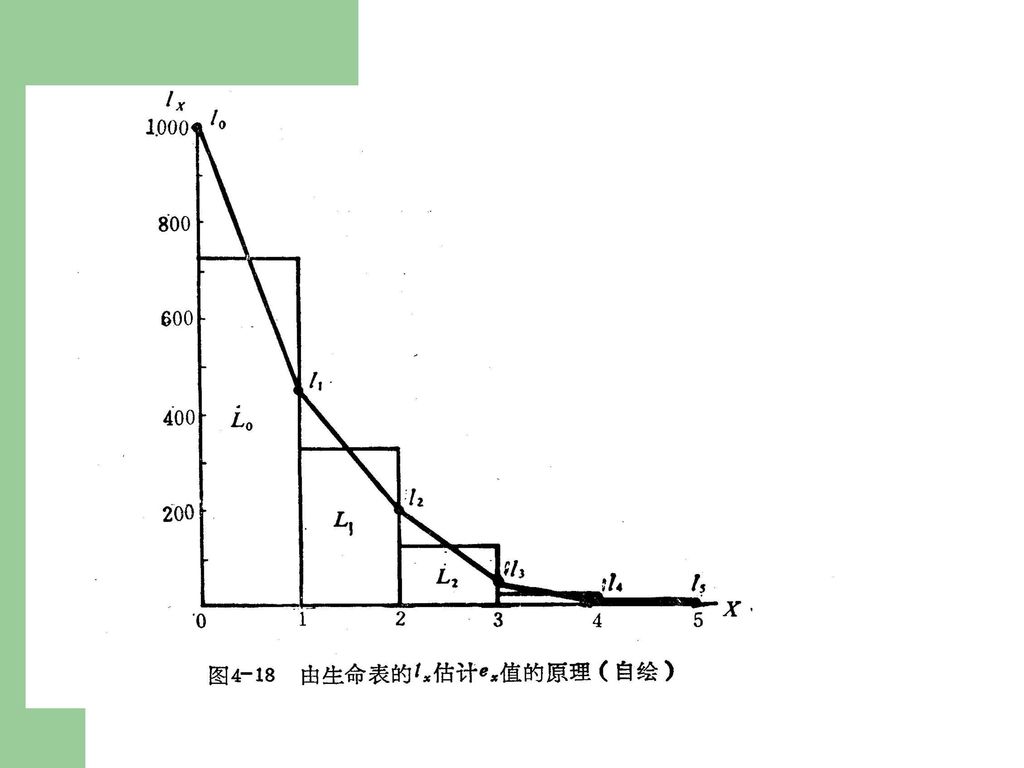
生命表有若干栏,即 X:为年龄期,即按年龄的分段,此表是以年来划分的,其它也可以按月﹑周或日来划分。X=0,表示1959年开始观察的那一年藤壶是0岁。 nx:为x期开始时的存活数。表示在每一年龄期开始时的存活藤壶数,它是通过实际观察而得到的。 lx :为存活率,它表示每一年龄期开始时的存活比例,它是按照下面公式计算的: lx = nx / n0 dx :为死亡数,它表示从X 到X + 1期的死亡数,即: dx = nx - nx+1 qx :为死亡率,它表示从X 到X + 1期的死亡率,即: qx = dx / nx Lx : 各年龄期中点的平均存活数,其值等于(nx + nx+1)/2 Tx : 种群全部个体的平均寿命总和,其值等于将生命表中各年龄期中点的平均存活数自下而上累加所得的值,如Tx = ∑Lx。 ex :x期开始时存活个体的平均期望寿命(生命期望),其值等于Tx / nx,如e1 = T1/ n1 = 224/142 = 1.58。显然,期望寿命计算要涉及到各年龄期中点的平均存活数(Lx)及其累加数(Tx)两项,它们与种群存活曲线的类型有关。
/2. Tx : 种群全部个体的平均寿命总和,其值等于将生命表中各年龄期中点的平均存活数自下而上累加所得的值,如Tx = ∑Lx。 ex :x期开始时存活个体的平均期望寿命(生命期望),其值等于Tx / nx,如e1 = T1/ n1 = 224/142 = 1.58。显然,期望寿命计算要涉及到各年龄期中点的平均存活数(Lx)及其累加数(Tx)两项,它们与种群存活曲线的类型有关。")
57
在编制生命表之前,首先要划分年龄阶段,如人常用5年,鱼类常用一年,枝角类常用1天等等。生命表中只要有nx或dx的观测值,就可以计算其他各栏,即:
nx+1 = nx – dx qx = dx/nx lx=nx/n0 但计算生命期望较复杂。首先求出每年龄期的平均存活数目=(nx + nx+1)/2,然后将这些结果列成表,再由表底向上累积Lx值得到Tx值,代表平均存活数与时间的乘积,即Tx = ΣLx。最后求出ex,即ex = Tx / nx,为平均生命期望,表示活到某年龄阶段的动物,平均还能活多少年的估计值。
/2,然后将这些结果列成表,再由表底向上累积Lx值得到Tx值,代表平均存活数与时间的乘积,即Tx = ΣLx。最后求出ex,即ex = Tx / nx,为平均生命期望,表示活到某年龄阶段的动物,平均还能活多少年的估计值。")
58
(二)存活曲线和期望寿命 1.存活曲线(survivorship curve)
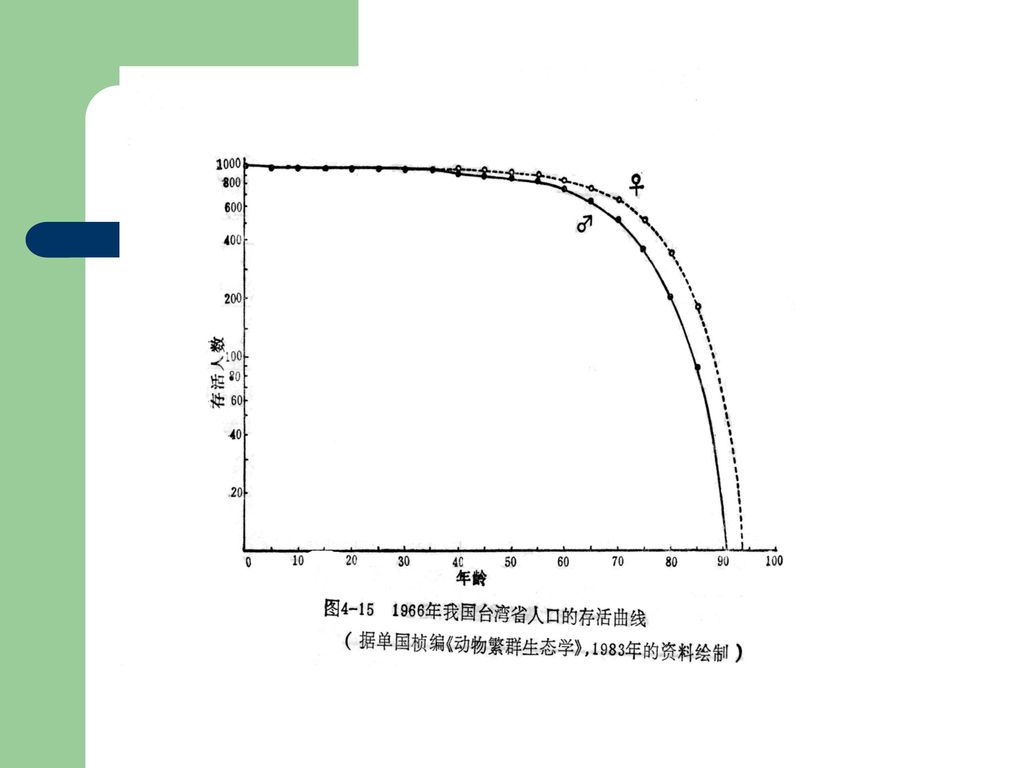
根据生命表可作存活曲线,存活曲线是以年龄为横坐标,存活个体数或存活率为纵坐标得到的曲线。表示随年龄增加而存活率(或死亡率)变化的情况。存活曲线中种群数量的数据是以对数标尺作图的。例如,下图表示人口的存活曲线。在50岁以下的人口死亡率很低,这与中世纪以前的人口存活曲线明显不同,它反映了医学和科学进展使得生态寿命延长,并接近于生理寿命的事实。同时,女人的平均寿命较男人的更长一些,这也是人口曲线的特点。
变化的情况。存活曲线中种群数量的数据是以对数标尺作图的。例如,下图表示人口的存活曲线。在50岁以下的人口死亡率很低,这与中世纪以前的人口存活曲线明显不同,它反映了医学和科学进展使得生态寿命延长,并接近于生理寿命的事实。同时,女人的平均寿命较男人的更长一些,这也是人口曲线的特点。")
61
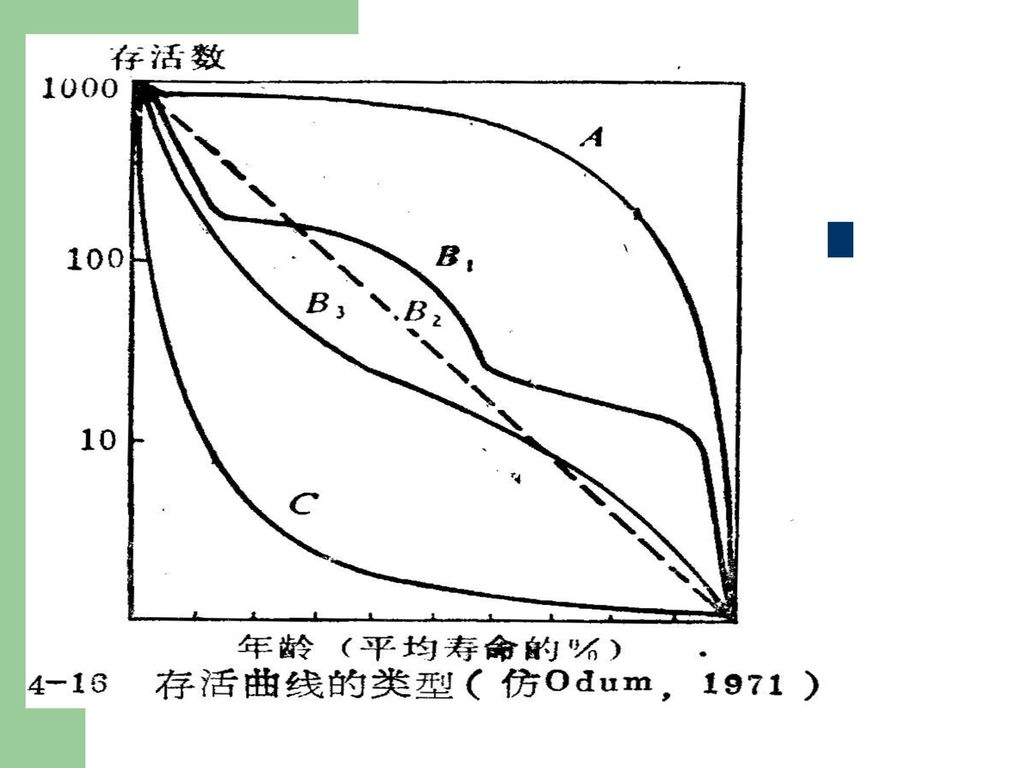
2.存活曲线的类型 存活曲线对研究种群的死亡过程是很有价值的。存活曲线对研究种群的死亡过程是很有价值的。Deevey(1947)以相对年龄(即以平均寿命的百分比表示的年龄,记作x)作为横坐标,存活率lx的对数作纵坐标,画成存活曲线,从而能够比较不同寿命的动物。 动物界中的存活曲线可划分为三个基本类型。 (1)A型,即凸型的存活曲线:表示种群接近于生理寿命之前,只有个别的死亡,即几乎所有个体都能达到生理寿命。如人类和大型哺乳动物。 (2)B 型,对角线型存活曲线:表示各年龄期的死亡率是相等的。大多数生物属此。如水螅、鸟类等。 (3)C型,凹型的存活曲线:幼体期死亡率很大,存活曲线骤然下降,但在度过此期以后,死亡率就低而稳定,如鱼类、牡蛎、寄生虫等属此。 现实生活中的生物种群,不会有完全这样典型的存活曲线,但可表现出接近某型或中间型。大多数生物居于A、B两型之间。
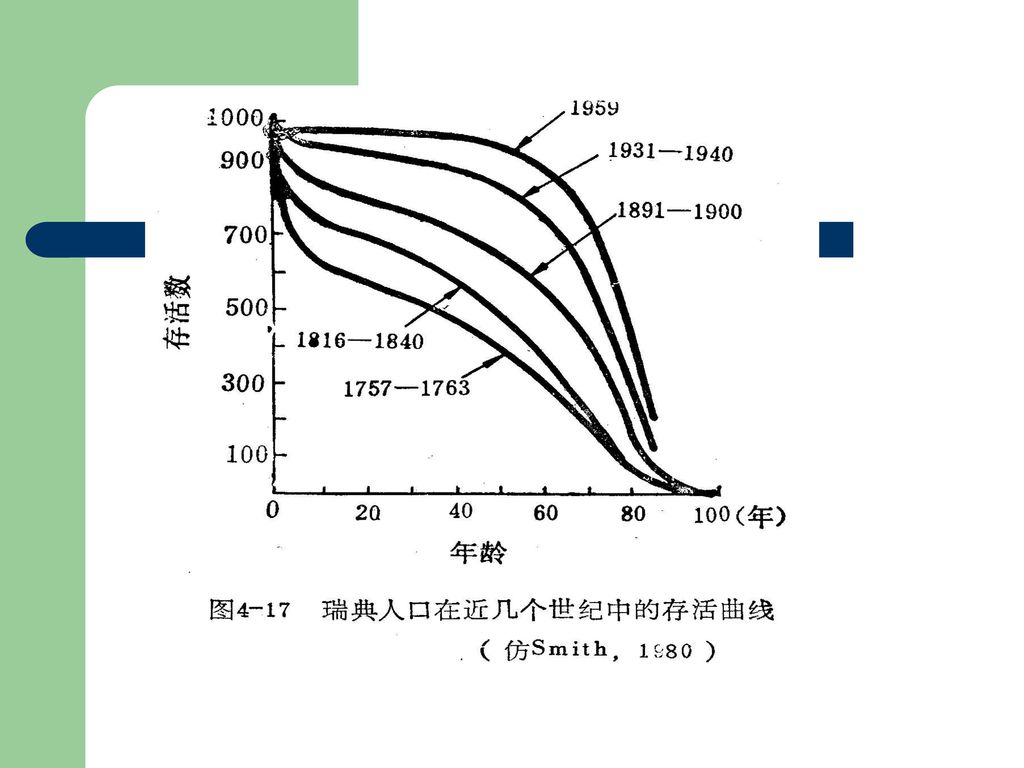
A型,即凸型的存活曲线:表示种群接近于生理寿命之前,只有个别的死亡,即几乎所有个体都能达到生理寿命。如人类和大型哺乳动物。 (2)B 型,对角线型存活曲线:表示各年龄期的死亡率是相等的。大多数生物属此。如水螅、鸟类等。 (3)C型,凹型的存活曲线:幼体期死亡率很大,存活曲线骤然下降,但在度过此期以后,死亡率就低而稳定,如鱼类、牡蛎、寄生虫等属此。 现实生活中的生物种群,不会有完全这样典型的存活曲线,但可表现出接近某型或中间型。大多数生物居于A、B两型之间。")
63
3.期望寿命 前已叙及,ex是指进入X龄期的个体,平均还能活多长时间的估计值,因此称之为生命期望或平均余生。

65
(三)特定年龄生育率(mx)和净生殖率(R0)
Lx mx栏是各年龄组的Lx与mx的乘积。将乘积相加,可求出总和∑Lx mx,称为净生殖率(R0)
")
66
(四)内禀增长能力 按照安德列沃斯和伯奇的定义,内禀增长力(innate capacity of increase)是:具有稳定年龄结构的种群,在食物和空间不受限制,同种其他个体的密度维持在最适水平,在环境中没有天敌,并在某一特定的温度、湿度、光照和食物条件下,种群的最大瞬时增长率,一般以rm表示。即rm是指在最适条件下种群的最大瞬时增长率(instantaneous rate)。种群的实际增长率是不断地变化着的。 rm 是一个种群的瞬时增长率。 前已叙及,内禀增长能力只是代表在特定条件下具有稳定年龄组配种群的最高瞬时增殖速率。 由于世代时间T 是一个近似的估算值,所以上述rm 也只能是一个近似值。
是:具有稳定年龄结构的种群,在食物和空间不受限制,同种其他个体的密度维持在最适水平,在环境中没有天敌,并在某一特定的温度、湿度、光照和食物条件下,种群的最大瞬时增长率,一般以rm表示。即rm是指在最适条件下种群的最大瞬时增长率(instantaneous rate)。种群的实际增长率是不断地变化着的。 rm 是一个种群的瞬时增长率。 前已叙及,内禀增长能力只是代表在特定条件下具有稳定年龄组配种群的最高瞬时增殖速率。 由于世代时间T 是一个近似的估算值,所以上述rm 也只能是一个近似值。")
67
rm 值精确值的计算方法,则要解如下方程式(Euler方程):
∞ ∑ e-rm﹒x lx mx = 1 x=0 这个方程式虽然很复杂,但可采用逐步逼近法求得精确值。 Euler方程的优点是,它把rm 直接与生命表中的lx 和mx 值联系了起来,只要有这两个值就可以直接求内禀增长能力,而且求出的值更加准确可靠。 内禀增长能力是一种瞬时增长率,它也可以转换为周限增长率(finite rate of increase)。其关系式是:λ= erm 。 一种生物的内禀增长能力决定于该种生物的生育力﹑寿命和发育速率。 由于内禀增长能力是种群增殖能力的一个重要综合指标,它不仅考虑到动物的出生率﹑死亡率,同时还将年龄结构﹑发育速度﹑世代时间等因素包括在内。因此,它可以敏感地反映出环境的细微变化,是特定种群对于环境质量反应的一个灵敏指标。
。其关系式是:λ= erm 。 一种生物的内禀增长能力决定于该种生物的生育力﹑寿命和发育速率。 由于内禀增长能力是种群增殖能力的一个重要综合指标,它不仅考虑到动物的出生率﹑死亡率,同时还将年龄结构﹑发育速度﹑世代时间等因素包括在内。因此,它可以敏感地反映出环境的细微变化,是特定种群对于环境质量反应的一个灵敏指标。")
68
§4 种群中的种内关系 1.个体间联络:通过视觉、听觉、触觉和化学信息(代谢产物—生物抑制和刺激作用;分泌物—促进集群、性行为、照顾后代,回归故乡等)。 2.直接斗争:同种相残,你死我活,而占领索饵场和栖息地处以及争夺异性的直接斗争不是你死我活的,通常是当一方失败即离开争斗场所,前者不置之死地而后快。 3.竞争:如食物的竞争,不同年龄、性别和状态的个体食谱差异可缓解之。 4. 互助:如集群、种内寄生。 5.领域制 6. 集群 7. 社会等级 8. 密度效应

69
动物的婚配制度 婚配制度是指种群内婚配的各种类型。婚配包括异性间相互识别、配偶的数目、配偶持续时间以及对后代的抚育等。因为雌配子大,雄配子小,所以每次婚配中雌性的投资大于雄性。雌性又可与雌性多次交配,所以雌性较雄性更关心交配的成功率。在高等动物婚配关系中,一般雌性是限制者,雄性常常不易接近雌性,是被限制者。由于这些原因,雄性常常因竞争雌性而发生格斗(如鹿),或建立吸引雌性的领域(如歌句鸟)等。这些特征就成为人们划分婚配制度亚类型的特征。一般认为高等动物最常见的婚配制度是一雄多雌制,而一雄一雌的单配偶制是原始的一雄多雌的多配偶制进化而来的。
,或建立吸引雌性的领域(如歌句鸟)等。这些特征就成为人们划分婚配制度亚类型的特征。一般认为高等动物最常见的婚配制度是一雄多雌制,而一雄一雌的单配偶制是原始的一雄多雌的多配偶制进化而来的。")
70
决定婚配制度类型的环境因素 决定动物婚配制度的主要生态因素可能是资源的分布,主要是食物和营巢地在空间和时间上的分布情况。例如,如果有一种食虫鸟,占据一片具有高质食物(如昆虫)资源并分布均匀的栖息地,雄鸟在其栖息地中各有其良好领域,那么雌鸟寻找没有配偶的雄鸟结成伴侣显然将比找已有配偶的雄鸟有利。因此,选择有利于形成一雄一雌制。不仅如此,若雄鸟也参与抚育雏鸟,单配偶也将比一雄多雌制有利。因为在一雄多雌制中,雄鸟要抚育多个雌鸟所产的雏鸟,这几乎是不可能的。再者,在一个领域中巢窝数越少、个体密度越低,也将给雏鸟提供更多食物,降低被捕食者捕食的机率。由此可见,高质而分布均匀的资源有利于产生一雄一雌的单配偶制。 相反,如果高质资源是呈斑点分布的,社群等级中处于高地位的雄鸟将选择并保卫资源最丰富的地方作为领域。在这种情况下,一旦占有资源丰富领域的雄鸟有了配偶以后,未有配偶的孤雌鸟选择配偶的困难会增加。此时,一雄二雌的多配偶制就产生了。 从高质领域到低质领域可视为一个连续的变化,在由高质到低质的变化过程中,单配偶制和多配偶制的相对利弊关系也随之相应变化。当达到从单配偶制转变到多配偶制的利弊相平衡的一点,可称为多配偶阈值(polygyny threshoid), 越过此阈值,多配偶制将比单配偶制更加有利。
资源并分布均匀的栖息地,雄鸟在其栖息地中各有其良好领域,那么雌鸟寻找没有配偶的雄鸟结成伴侣显然将比找已有配偶的雄鸟有利。因此,选择有利于形成一雄一雌制。不仅如此,若雄鸟也参与抚育雏鸟,单配偶也将比一雄多雌制有利。因为在一雄多雌制中,雄鸟要抚育多个雌鸟所产的雏鸟,这几乎是不可能的。再者,在一个领域中巢窝数越少、个体密度越低,也将给雏鸟提供更多食物,降低被捕食者捕食的机率。由此可见,高质而分布均匀的资源有利于产生一雄一雌的单配偶制。 相反,如果高质资源是呈斑点分布的,社群等级中处于高地位的雄鸟将选择并保卫资源最丰富的地方作为领域。在这种情况下,一旦占有资源丰富领域的雄鸟有了配偶以后,未有配偶的孤雌鸟选择配偶的困难会增加。此时,一雄二雌的多配偶制就产生了。 从高质领域到低质领域可视为一个连续的变化,在由高质到低质的变化过程中,单配偶制和多配偶制的相对利弊关系也随之相应变化。当达到从单配偶制转变到多配偶制的利弊相平衡的一点,可称为多配偶阈值(polygyny threshoid), 越过此阈值,多配偶制将比单配偶制更加有利。")
71
婚配制度的类型 按配偶数有单配偶制(monogamy)和多配偶制(polygamy),后者又分一雄多雌制(polygyny)和一雌多雄制(polyandry)。 ①一雄多雌制:最常见,例如海狗营集群生活,尤其在繁殖期内。通常雄兽首先到达繁殖基地(如岩岸,沙滩或大块浮冰等),争夺并保护领地。雌兽到达较晚,不久就生育幼仔。一只雄兽独占雌兽少则3只,多至40只以上。 ②一雌多雄制:与一雄多雌制比较,动物中一雌多雄是很稀见的,据统计,鸟类中有1%,如距翅水雉(Jacana spinosa)。 ③单配偶制:动物中单配偶制是比较少见的,只有鸟类以一雄一雌制较普遍,例如鸳鸯、天鹅、单顶鹤和许多鸣禽。哺乳类动物中单配偶制是一个例外,例如狐、鼬和河狸。
,争夺并保护领地。雌兽到达较晚,不久就生育幼仔。一只雄兽独占雌兽少则3只,多至40只以上。 ②一雌多雄制:与一雄多雌制比较,动物中一雌多雄是很稀见的,据统计,鸟类中有1%,如距翅水雉(Jacana spinosa)。 ③单配偶制:动物中单配偶制是比较少见的,只有鸟类以一雄一雌制较普遍,例如鸳鸯、天鹅、单顶鹤和许多鸣禽。哺乳类动物中单配偶制是一个例外,例如狐、鼬和河狸。")
72
领域制 领域制(territory)是指某种动物的个体或特定群体(配偶或家族)能防卫的不让同种其他成员侵入的地域。在这种情况下,防卫的对方,通常是指同种的其他个体或群体。蜘蛛虽然攻击接近于身边的所有动物,但这不能称为领域。领域的含义是指同种内的社会关系。如香鱼具领域制,香鱼对侵入到他们范围的其他鱼类不怎么关心,如同种动物进入狭窄的空间,则通过斗争就形成攻击等级。但是,把领域作为其社会的基本行动方式确定下来,在这样的动物中,即使侵入者的等级高,只要是其差别不很大,领域所有者通常是会取胜的。这样领域就能保障该种的一定个体或群体有一个最小的空间。
是指某种动物的个体或特定群体(配偶或家族)能防卫的不让同种其他成员侵入的地域。在这种情况下,防卫的对方,通常是指同种的其他个体或群体。蜘蛛虽然攻击接近于身边的所有动物,但这不能称为领域。领域的含义是指同种内的社会关系。如香鱼具领域制,香鱼对侵入到他们范围的其他鱼类不怎么关心,如同种动物进入狭窄的空间,则通过斗争就形成攻击等级。但是,把领域作为其社会的基本行动方式确定下来,在这样的动物中,即使侵入者的等级高,只要是其差别不很大,领域所有者通常是会取胜的。这样领域就能保障该种的一定个体或群体有一个最小的空间。")
73
领域制结论 保卫领域的方式很多,例如,以鸣叫、气味标志或特异的姿势向入侵者宣告具领主的领域范围;以威胁或直接进攻驱赶入侵者,称为领域行为。具领域性种类在脊椎动物中最多,尤其是鸟兽,某些昆虫也有之。 动物领域行为的规律性结论:领域面积随领域占有者的体重而扩大,领域的大小必须必须以能保证供应足够的食物资源为前提,动物越大,需要资源越多,领域面积也就越大。领域面积受食物质量的影响,食肉动物的领域面积较同样体重的食草动物大,并且体重越大,这种差别也越大。其原因是食肉动物获取食物需要消耗更多的能量,包括追击和捕杀。领域行为和面积往往随生活史,尤其是繁殖节律而变化。

74
领域制的生态意义 ①由于接近特定的场所,营巢地,容易逃避敌害。 ②使种群分散,保证食物资源,预防因拥挤而造成坏影响。
③有利于雌雄的形成和保持。 容易保护幼体。

75
集群 动物集群生活的生态意义在于 (1)拥挤效应:动物集群时繁殖过剩和种群密度过高所产生的有害效应称为拥挤效应。如食物空间的竞争加剧,排泄物增多,产生心理和生理压力等。 (2)集群效应:同一种动物在一起生活所产生的有利作用称为集群效应。 生物产生集群的原因 ① 对栖息地食物、光、温、水等生态因子的共同需要; ② 对昼夜天气或季节气温的共同反应; ③ 繁殖的结果; ④ 被动运送的结果; 由于个体之间社会吸引力(social attraction)相互吸引的结果。动物群体由环境因素所决定的称为集会,由社会吸引力引起的称为社会。 (3)最小种群原则:对于一些集群生活的动物种类,如果数量太小,低于集群的临界下限,则该种群就不能正常生活,甚至不能生存。这就是所谓的最小种群原则。
相互吸引的结果。动物群体由环境因素所决定的称为集会,由社会吸引力引起的称为社会。 (3)最小种群原则:对于一些集群生活的动物种类,如果数量太小,低于集群的临界下限,则该种群就不能正常生活,甚至不能生存。这就是所谓的最小种群原则。")
76
集群的生态意义 集群的生态学意义 ① 集群有利于提高捕食效率;
① 集群有利于提高捕食效率; ② 集群有利于共同防御敌害,共同警戒,分工协作,共同逃离或混乱效应和共同防御敌害。 ③ 改变小生境,如越冬鲤鱼的集群减少散热,蜜蜂冬季肌肉颤抖加热升温,夏季运水煽翼降温,40℃降到36℃。 ④有利于某些动物提高学习效率,增加学习机会,取长补短。 ⑤促进繁殖,利于求偶、交配、产仔和育幼。

77
社会等级 社会等级现象是社会组织的特征之一。在许多动物群体当中,动物各自有一定的等级地位,等级地位较高的优势个体比等级地位较抵的从属个体优先获得资源,满足其食物、栖息场所、配偶等需要。动物种群中各个个体之间的这种优势顺序就称为社会等级(social hierarchys)或优势顺序( dominance order)。社会等级形成的基础是优势行为(dominance behaviour)。
或优势顺序( dominance order)。社会等级形成的基础是优势行为(dominance behaviour)。")
78
社会等级的类型 ①一长式(despotic hierarchy)或独霸式。社群当中的所有个体只受一个优势个体所支配,其它个体之间没有等级差别。如猴。 ②单线式(linear hierarchy)或系列式。群体当中的每个个体都有一定的社会地位,甲支配乙,乙支配丙,丙支配丁……。如家鸡。 ③循环式(cyclic 或tridic hierarchy)或三角式。甲支配乙,乙支配丙,而乙又支配甲。这种等级制不因个体力量、外形和体重的大小以及是否伤残而受影响。见于多雄体的灵长类,它们往往形成复杂的三角等级关系和年龄等级互相配合的支配系统。主雄和次雄之间多有互助现象,如狒狒和黑猩猩等。
或三角式。甲支配乙,乙支配丙,而乙又支配甲。这种等级制不因个体力量、外形和体重的大小以及是否伤残而受影响。见于多雄体的灵长类,它们往往形成复杂的三角等级关系和年龄等级互相配合的支配系统。主雄和次雄之间多有互助现象,如狒狒和黑猩猩等。")
79
社会等级的特点 ①排他性:外来者低级或被驱逐;
②社会惰性:等级是通过相互接触或格斗形成的,一段时间不在发生地位争夺,相遇时从属个体顺从的姿态或离开。 ③权利欲:改朝换代; ④雌雄等级制互相分开; ⑤优势个体的利他性:优势个体牺牲自我利益而增加其他个体获得利益的行为称为利他行为( altruism)。
。")
80
社会等级的生态意义 ① 非争斗性地获得有限资源; ②调节种群数量。

81
密度效应 密度效应(density effect)在一定时间内,当种群的个体数目增加时,就必定会出现邻接个体之间的相互影响,或称为邻接效应(the effect of neighbours)。 (1)最后产量恒值法则 (2)-3/2自疏法则
-3/2自疏法则.")
82
(1)最后产量恒值法则 在一定范围内,当条件相同时,不管一个种群的密度如何,最后产量差不多总是一样的。这就是最后产量恒定法则。即
Y=W.d=Ki 式中:W——植物个体平均重量; d ——密度; Y——单位面积产量; Ki——常数。 最后产量恒值法则的原因是在高密度情况下植株之间的光、水、营养物的竞争十分激烈,在有限的资源中,植株的生长率降低,个体变小。

83
( 2)-3/2自疏法则 随着播种密度的提高,种内对资源的竞争不仅影响到植株生长发育的速度,也影响到植株的成活率。在高密度的样方中,有些植株死亡了,于是种群开始出现“自疏现象”(self-thinning)。日本学者Yoda等(1963)把自疏过程中存活个体的平均株干重(W)与种群密度(d)之间的关系用下式来表示:W=Cd-a 两边取对数得:lgW=lgC-algd 英国生态学家J.L.Harper(1981)等对黑麦草(Lolium perenne)的研究发现a为一个恒值等于2/3。因此被称为-3/2自疏法则。
。日本学者Yoda等(1963)把自疏过程中存活个体的平均株干重(W)与种群密度(d)之间的关系用下式来表示:W=Cd-a. 两边取对数得:lgW=lgC-algd. 英国生态学家J.L.Harper(1981)等对黑麦草(Lolium perenne)的研究发现a为一个恒值等于2/3。因此被称为-3/2自疏法则。")
84
§5 种群的增长 种群生态学历来是最为活跃的领域。其中,种群数量动态是种群研究的核心内容。种群的数量变动决定于出生率、死亡率、迁入和迁出,在一般情况下都假定迁入和迁出是平衡的。这样,种群的增长主要取决于出生率和死亡率的对比关系。在单位时间内,出生率与死亡率之差为增长率,也就是单位时间内种群数量增加的百分数。种群增长率可用单位时间内个体数或生物量的绝对值表示,也可以用增长量对原有量的百分比表示。 在自然条件下,任何生物的种群与生物群落中的其他生物密切相关,不能从其中孤立开来。因此,严格地说,单种种群只有在实验室内才有可能存在。但是人们为了要了解种群的增长和动态规律,往往从研究分析单种种群开始。

85
一个物种,从其进入新的栖息地,经过增长并建立起种群以后,一般有如下几种可能:种群增长J,S型,季节消长。
较长期地维持在几乎同一水平上,称为种群平衡(population equilibrium)。 经受不规则的波动或起落(irregular flunctuation) 规则的或周期性的波动(regular or cyclic flunctuation),有时也称振荡(oscillation)。 种群衰落(population decline) 种群灭亡(population extinction) 有时种群数量在短期中迅速增长,称为种群大发生或暴发(population outbreak),如海洋中的赤朝现象。又如槐叶萍1952年在澳大利亚爆发。 在种群大发生后,往往出现大批死亡,种群数量剧烈下降,即种群崩溃(population crash)。
。 经受不规则的波动或起落(irregular flunctuation) 规则的或周期性的波动(regular or cyclic flunctuation),有时也称振荡(oscillation)。 种群衰落(population decline) 种群灭亡(population extinction) 有时种群数量在短期中迅速增长,称为种群大发生或暴发(population outbreak),如海洋中的赤朝现象。又如槐叶萍1952年在澳大利亚爆发。 在种群大发生后,往往出现大批死亡,种群数量剧烈下降,即种群崩溃(population crash)。")
86
生态入侵 由于人类有意识或无意识地把某种生物带入适宜其栖息和繁衍的地区,种群不断扩大,分布区逐步稳定地扩展,这种过程称生态入侵(ecological invasion)。 (1)欧洲穴兔——16年在澳大利亚推进1770km——与牛羊争草——黏液瘤病毒杀之。 (2)克氏螯虾 (3)福寿螺 (4)多形饰贝
克氏螯虾. (3)福寿螺. (4)多形饰贝.")
87
野外研究较困难,而实验室种群研究较多,因为条件易保持稳定。在实验种群的研究中,往往是先提出假设,建立模型,然后加以检验,这种模型常为数学模型,通过此模型可理解各种生物和非生物因子的相互作用如何影响各种生物种群的动态。因此,我们感兴趣的不是特定公式的数学细节,而是模型的结构及模型中各个量的生物学意义。 种群生长,各种生物不同,但按其变动曲线可以分为“J”和“S”型两中基本类型。两者之间存在着各种中间类型。 在自然条件下,对于现实种群来说,不可能有持久的无限制增长,因为环境是有限的,环境中的空间﹑食物﹑气候﹑天敌等条件都不可能无限制地供养种群无止境的增长。但是,我们如果假定有一个理想种群,处于某一个无限制的环境中,那么有关它可能怎样增长的情况,对于进一步分析种群在有限环境中的增长是很有用的。理想种群包含如下四个假设:①种群在无限的环境中增长,不受资源﹑空间等条件的限制;②世代不相重叠,增长是不连续的,或称离散的;增长倍数相同,出生率与死亡率稳定;③种群没有迁入和迁出;④种群没有年龄结构。因此,我们将从无限环境中的理想种群增长的研究入手逐渐加以深化。

88
一﹑种群在无限环境中的指数式增长—J型生长曲线
(一)世代不相重叠种群的离散增长模型(差分方程) (二) 世代重叠种群的连续增长模型(微分方程)
世代不相重叠种群的离散增长模型(差分方程) (二) 世代重叠种群的连续增长模型(微分方程)")
89
(一)世代不相重叠种群的离散增长模型(差分方程)
假定有一种动物,一年只繁殖一次,寿命也只有一年,那么每年的世代就不会相互重叠。 单种种群的数量,理论上决定于出生率、死亡率和种群的起始数量。种群数量增长的方程式概括起来即Nt = N0 λt= 20×10t。这个公式就代表世代不重叠的种群在无限环境中的增长过程,我们可以称为离散种群在无限环境中增长的数学模型。 在上述方程中有两个变量:N 、t,一个常数λ。N 表示种群的数量,t表示时间。λ表示N 与t的函数关系,称为周限增长率(finite rate of increase)。周限增长率(λ) 表示种群以每年(或其他时间单位)为前一年以Nt+1/Nt倍的速率而增长的增长率。这种增长形式称为几何级数式增长或指数式增长。以t为横坐标,Nt为纵坐标作图,曲线呈“J”型,所以指数式增长模型又称“J”型增长模型。
。周限增长率(λ) 表示种群以每年(或其他时间单位)为前一年以Nt+1/Nt倍的速率而增长的增长率。这种增长形式称为几何级数式增长或指数式增长。以t为横坐标,Nt为纵坐标作图,曲线呈 J 型,所以指数式增长模型又称 J 型增长模型。")
90
(二) 世代重叠种群的连续增长模型(微分方程)
连续生殖种群是指在任一时间内,这个种群都存在着年龄不同的个体,总有新的个体产生,具体体现在体长——体重参差不齐,既难弄清楚种群中究竟有多少个同龄子代,又无法追踪其存活过程。亦即生物的世代是相互重叠的,一般细菌、藻类、浮游植物、浮游动物属此。这样的种群,出生和死亡都是连续的、不间断的, 所以种群增长是一个连续的过程。对于世代重叠种群的连续增长过程,通常用微分方程来描述。即: dN/ dt = rN

91
推导过程1 在一个孤立的生态系统中,种群数量(N)随时间的推移而变动,变动量是出生数(B)+迁入数(I)和死亡数(D)+迁出数(E)的差数,既 dN/dt = B + I – D - E 假设I=E,则 dN/dt = B – D 由于出生数和事物数与种群密度(N)成比例,所以B = bN(出生数=出生率×种群密度);D = dN(死亡数=死亡率×种群密度),那么: dN/dt = B – D = bN – dN =N(b-d) = rN dN/dt 表示在时间t 瞬间的变化中,种群数量N 的变化率。
成比例,所以B = bN(出生数=出生率×种群密度);D = dN(死亡数=死亡率×种群密度),那么: dN/dt = B – D = bN – dN =N(b-d) = rN. dN/dt 表示在时间t 瞬间的变化中,种群数量N 的变化率。")
92
推导过程2 表示种群中平均每个个体瞬时的变化情况。其积分式为: Nt = N0 e rt
其中N0 、Nt 、t的定义如前;e = ……,称为指数常数,即自然对数的底。r是种群的瞬时增长率。瞬时增长率(r) 表示特定环境条件下种群瞬时出生率和瞬时死亡率之差。即r是描述种群在无限环境中中呈几何级数增长的瞬时增长能力的。种群的内禀增长能力r m就是种群在最适条件下的最大瞬时增长率。b和d 表示种群的瞬时出生率和瞬时死亡率,那么瞬时增长率r就等于(b - d),即r =b – d(假设没有迁入和迁出)。
 表示特定环境条件下种群瞬时出生率和瞬时死亡率之差。即r是描述种群在无限环境中中呈几何级数增长的瞬时增长能力的。种群的内禀增长能力r m就是种群在最适条件下的最大瞬时增长率。b和d 表示种群的瞬时出生率和瞬时死亡率,那么瞬时增长率r就等于(b - d),即r =b – d(假设没有迁入和迁出)。")
93
种群的瞬时增长率r 描述了种群在无限环境中呈几何级数式增长的瞬时增长能力。因此,前述的内禀增长能力rm 就是一种种群的瞬时增长率。实际上,rm 是指rmaximum,即最大的r,。但r 的含义更广泛,而rm 是指特定条件下的最大r 。 无论是周限增长率或是瞬时增长率,它们所表示的生物学含义都有四种情况,其间的对应关系如下:

94
一般说来,在种群动态研究中,种群瞬时增长率较周限增长率应用更为广泛,这是因为:
(1)r为正值、负值或零,能表示种群的正增长;负增长和零增长;而λ表示的是t + 1时和t时的种群Nt + 1和Nt的比率。例如,当有两个种群,它们的周限增长率分别为λ1= 1.649, λ2= 0.607,这两个数值只能说明前一个种群是上升的,后一个种群是下降的,但看不出增加的速度和下降速度的关系。如果把它们变换为瞬时增长率,r1 = ln = 0.5, r2 = ln = -0.5,就能看出前一种群的增长率正好等于后一种群的减少率。由于瞬时增长率是表明种群连续变化的情况,因此在计算时可以根据需要把单位时间作无限的分割。 (2)r具有可加性。如中华盒形藻的瞬时增长率rm = 是以天为时间单位表示,若以周为时间单位,则rm = ×7 = 。因此,种群的瞬时增长率r是种群动态研究中一个十分有用的参数。
r为正值、负值或零,能表示种群的正增长;负增长和零增长;而λ表示的是t + 1时和t时的种群Nt + 1和Nt的比率。例如,当有两个种群,它们的周限增长率分别为λ1= 1.649, λ2= 0.607,这两个数值只能说明前一个种群是上升的,后一个种群是下降的,但看不出增加的速度和下降速度的关系。如果把它们变换为瞬时增长率,r1 = ln = 0.5, r2 = ln = -0.5,就能看出前一种群的增长率正好等于后一种群的减少率。由于瞬时增长率是表明种群连续变化的情况,因此在计算时可以根据需要把单位时间作无限的分割。 (2)r具有可加性。如中华盒形藻的瞬时增长率rm = 是以天为时间单位表示,若以周为时间单位,则rm = ×7 = 。因此,种群的瞬时增长率r是种群动态研究中一个十分有用的参数。")
95
二﹑种群在有限环境中逻辑斯谛增长—“S”型增长
(一)世代不相重叠种群 (二)连续生殖种群
世代不相重叠种群. (二)连续生殖种群.")
96
(一)世代不相重叠种群 无限环境中的差分方程Nt+1=λNt 描述了非连续生殖种群增长的适当模型。在此方程式中,周限增长率λ与种群密度无关。但是大多数水生生物种群的增长率随密度而改变,因此必须考虑密度改变对种群增长率的影响问题。最简单的模型是假定密度与增长率关系是线性的。λ随密度增加而下降,假设种群平衡密度Neq为100个个体(即此时λ= 1),由此平衡密度每偏离一个个体,增长率λ减少或增加2%(即0.02),亦即λ随N值而改变,其关系式为: λ = 1.0 – B ( Nt – Neq) 种群增长模型为: N t+1 =λNt = [1.0 – B ( Nt – Neq)]Nt
,由此平衡密度每偏离一个个体,增长率λ减少或增加2%(即0.02),亦即λ随N值而改变,其关系式为: λ = 1.0 – B ( Nt – Neq) 种群增长模型为: N t+1 =λNt = [1.0 – B ( Nt – Neq)]Nt.")
98
关于这个模型的行为,可以用改变参数B值大小的方法实验出来.
(1)如设B=0.011, Neq = 100,种群初时为10个个体,即N0=10,那么 N1 =[ (10-100)]10 = (1.99)(10) = 19.9 N2 =[ ( )]19.9 = (1.881)(19.9) = 37.4 N3 = 63.1, N4 = 88.7, N5 = 99.7 结果表明,种群密度平滑地趋向于平衡点100. (2) 如设B= 0.025, Neq = 100,种群初时为10个个体,即N0=50,那么 N1 =[ (50-100)]50 = (2.23)(50) = 112.5 N2 =[ ( )]112.5 = (0.6875)(112.5) = 77.34 N3 = , N4 = 55.80, N5 = 117.5 结果说明,种群稳定地振荡(0scillation)。
如设B=0.011, Neq = 100,种群初时为10个个体,即N0=10,那么. N1 =[ (10-100)]10 = (1.99)(10) = N2 =[ ( )]19.9 = (1.881)(19.9) = N3 = 63.1, N4 = 88.7, N5 = 结果表明,种群密度平滑地趋向于平衡点100. (2) 如设B= 0.025, Neq = 100,种群初时为10个个体,即N0=50,那么. N1 =[ (50-100)]50 = (2.23)(50) = N2 =[ ( )]112.5 = (0.6875)(112.5) = N3 = , N4 = 55.80, N5 = 结果说明,种群稳定地振荡(0scillation)。")
99
(3)如N0=10, Neq = 100,而B分别为0. 013,0. 023. 033时,我们从图中可以看到,当B=0
(3)如N0=10, Neq = 100,而B分别为0.013, 时,我们从图中可以看到,当B=0.013时,种群平滑地向平衡点上升,并出现稳定的种群;当B=0.023 时,种群稳定地振荡,其周期为2个世代,当B=0.033时,种群表现出增加振幅的振荡,并于第8世代消亡。 此模型行为的不同取决于B值的大小,即种群增长率随密度增减而改变其速度。
如N0=10, Neq = 100,而B分别为0.013, 时,我们从图中可以看到,当B=0.013时,种群平滑地向平衡点上升,并出现稳定的种群;当B=0.023 时,种群稳定地振荡,其周期为2个世代,当B=0.033时,种群表现出增加振幅的振荡,并于第8世代消亡。 此模型行为的不同取决于B值的大小,即种群增长率随密度增减而改变其速度。")
100
(二)连续生殖种群 指数增长模型所描述的数量增长是没有界限的。但是在自然界中,甚至在实验室内,种群的增长不可能不受限制。虽然许多动、植物物种群在实验室条件下或野外的自然状态下,在其生活史的某一阶段可表现出几何级数的增长过程,但这种按指数增长的内禀增长能力只能在短期中出现。随着种群密度的上升,食物、空间等资源需要量迅速地增加,环境条件的限制和种内个体间竞争的加剧,必然影响种群的出生率和存活率,表现为当种群数量增加到一定限度后,种群增长率逐渐降低,最后达到出生率与死亡率相等,甚至出现负增长。总之,无限的“J”型增长是暂时的,而有限的增长是必然的。

101
为了使理想种群现实化,将以指数增长模型为基础,加进了限制无限增长的修正项,将其代入指数方程,得:
dN /dt = rN (1 – N/K ) = rN ( K - N)/K 其中,N ,t ,r的定义和指数增长方程相同。但增加了一个修正项(1 – N/K )称为阻滞系数,环境阻力、剩余空间(residual space)或称未利用的生长机会(unutilized opportunity for growth)。 K为环境负载量、环境负荷量以及环境容纳量(enviromental carring capacity)。指一个特定种群在一个时期内在特定的环境条件下生态系统所支持的种群有限大小。即种群增长的最高水平。K与环境有关,包括空间、食物和生物物理因素。由于环境的不稳定性,实际中容纳量并非是一个常数。
 = rN ( K - N)/K. 其中,N ,t ,r的定义和指数增长方程相同。但增加了一个修正项(1 – N/K )称为阻滞系数,环境阻力、剩余空间(residual space)或称未利用的生长机会(unutilized opportunity for growth)。 K为环境负载量、环境负荷量以及环境容纳量(enviromental carring capacity)。指一个特定种群在一个时期内在特定的环境条件下生态系统所支持的种群有限大小。即种群增长的最高水平。K与环境有关,包括空间、食物和生物物理因素。由于环境的不稳定性,实际中容纳量并非是一个常数。")
102
对(1 – N/K )这个修正项,可以作如下分析:
1.当N趋向于零时,(1 – N/K )就逼近于1,这表示K 空间几乎尚未被利用,rN能100%的实现,种群接近于指数增长,或种群潜在的最大增长率能完全的实现。 2.如果N趋向于K时,于是(1 – N/K )就逼近于零,这表示K 空间几乎全部被利用,rN所描述的种群可能有的最大增长率完全不能实现,种群增长率等于零。 3.在这两种情况之间,当种群数量由小逐渐增大到K值时,(1 – N/K )项就由1逐渐变小而到零,这表示可供种群继续增长的“剩余空间”越来越小,rN所描述的种群最大增长率可以实现的程度就越来越小。 上式就是著名的逻辑斯谛种群增长模型(Logistic equation )。
就逼近于1,这表示K 空间几乎尚未被利用,rN能100%的实现,种群接近于指数增长,或种群潜在的最大增长率能完全的实现。 2.如果N趋向于K时,于是(1 – N/K )就逼近于零,这表示K 空间几乎全部被利用,rN所描述的种群可能有的最大增长率完全不能实现,种群增长率等于零。 3.在这两种情况之间,当种群数量由小逐渐增大到K值时,(1 – N/K )项就由1逐渐变小而到零,这表示可供种群继续增长的 剩余空间 越来越小,rN所描述的种群最大增长率可以实现的程度就越来越小。 上式就是著名的逻辑斯谛种群增长模型(Logistic equation )。")
103
种群增长率= (种群潜在的最大增长)* (最大增长之可实现程度)
其积分式为 N = K / ( 1 + e a - rt) 其中a =r/K, 其值取决于N0, 是表示曲线对原点的相对位置。逻辑斯谛方程所配合的曲线称为逻辑斯谛曲线(logistic curve),因其形状呈“S”型故也常称之为S型曲线。环境阻力(environmental resistance)( K - N)/K即是“J”型曲线和“S”型曲线之间的部分(图8-12)。总之,逻辑斯谛曲线是一条向着环境负荷量(K)逼近的“S”型增长曲线 “语言文字模型: 种群增长率= (种群潜在的最大增长)* (最大增长之可实现程度)
 其中a =r/K, 其值取决于N0, 是表示曲线对原点的相对位置。逻辑斯谛方程所配合的曲线称为逻辑斯谛曲线(logistic curve),因其形状呈 S 型故也常称之为S型曲线。环境阻力(environmental resistance)( K - N)/K即是 J 型曲线和 S 型曲线之间的部分(图8-12)。总之,逻辑斯谛曲线是一条向着环境负荷量(K)逼近的 S 型增长曲线. 语言文字模型: 种群增长率= (种群潜在的最大增长)* (最大增长之可实现程度)")
104
“S”和“J”型曲线外观上有两点主要区别:

105
由指数增长模型发展到逻辑斯谛增长模型,从参数讲,只增加一个K值,从方程式来说,也只增加一个( K - N)/K项,但它阐明的实质内容则是生态学中很重要的机制:具体体现在:
(2)当种群密度上升时,种群能实现的有效增长率逐渐降低,即种群增长具有密度效应。这就是说,在种群密度与增长率之间,存在着反馈机制。一方面,随着时间,密度在不断上升;另一方面,随着密度上升,种群的增长率随之下降。显然,这是一种十分明显的密度制约作用。 (3)密度增长对于增长率的下降的抑制效应是按比例的,即每增加一个个体,就产生1/K的影响。因此,种群能实现的增长程度为种群密度本身的函数,即(1-N/K). (4) 逻辑斯谛增长方程所描述的增长曲线是呈“S”型的。“S”型的逻辑斯谛曲线在N = K /2处有一个拐点,在拐点上dN /dt最大。在达到拐点前,dN /dt随种群增加而上升,称为正加速期;在拐点之后,dN /dt随种群增加而下降,称为负加速期。
当种群密度上升时,种群能实现的有效增长率逐渐降低,即种群增长具有密度效应。这就是说,在种群密度与增长率之间,存在着反馈机制。一方面,随着时间,密度在不断上升;另一方面,随着密度上升,种群的增长率随之下降。显然,这是一种十分明显的密度制约作用。 (3)密度增长对于增长率的下降的抑制效应是按比例的,即每增加一个个体,就产生1/K的影响。因此,种群能实现的增长程度为种群密度本身的函数,即(1-N/K). (4) 逻辑斯谛增长方程所描述的增长曲线是呈 S 型的。 S 型的逻辑斯谛曲线在N = K /2处有一个拐点,在拐点上dN /dt最大。在达到拐点前,dN /dt随种群增加而上升,称为正加速期;在拐点之后,dN /dt随种群增加而下降,称为负加速期。")
106
种群生长的几个阶段: 逻辑斯谛增长是最常见的生长形式,可以说是种群增长的共性,并称之为种群增长模型。可划分为 下列几个阶段(如图8-13)
增长期:种群数量增加,K-N>0 诱发期:数量暂时无明显增长或增长极慢; 指数增长期:增长率接近种群的内禀增长率。呈指数式增长。 直线增长期:数量继续增加,但增长率和增长量逐渐下降。 平衡期:数量达到高峰后保持稳定的阶段。但也有颤动或波动。 消落期:随着死亡率的增大,种群数量逐渐下降的阶段。初期指数下降,以后非指数下降。 以上对藻类、细菌或动物等的培养有重要指导意义。

107
三、具有时滞的种群增长模型 (一)具有时滞的种群离散增长模型 (二)具有时滞的种群连续增长模型
具有时滞的种群离散增长模型 (二)具有时滞的种群连续增长模型")
108
(一)具有时滞的种群离散增长模型 由于种群密度增加而带来的降低种群增长的效应是有时滞的,即高密度对于出生率的影响往往出现在经过较长的时间以后。在某些昆虫中,高密度对出生率的影响要延迟到成虫种群减少时才表现出来,这大约要经过一个世代的时间。因此,在非连续生殖种群中,在考虑时滞的增长模型时,一般假定t世代的种群增长率,不是依赖于t世代的密度,而是依赖于t-1世代的密度。

109
最简单的具有时滞的种群离散增长模型 N t+1 =λNt = [1.0 – B ( N t-1 – Neq)]Nt
这与无时滞的种群离散增长模型相似,只不过– B ( Nt – Neq)改为– B ( N t-1 – Neq),即种群前一代密度N t-1影响了种群的增长率。
![最简单的具有时滞的种群离散增长模型 N t+1 =λNt = [1.0 – B ( N t-1 – Neq)]Nt](http://slidesplayer.com/slide/11259827/61/images/109/%E6%9C%80%E7%AE%80%E5%8D%95%E7%9A%84%E5%85%B7%E6%9C%89%E6%97%B6%E6%BB%9E%E7%9A%84%E7%A7%8D%E7%BE%A4%E7%A6%BB%E6%95%A3%E5%A2%9E%E9%95%BF%E6%A8%A1%E5%9E%8B+N+t%2B1+%3D%CE%BBNt+%3D+%5B1.0+%E2%80%93+B+%28+N+t-1+%E2%80%93+Neq%29%5DNt.jpg "这与无时滞的种群离散增长模型相似,只不过– B ( Nt – Neq)改为– B ( N t-1 – Neq),即种群前一代密度N t-1影响了种群的增长率。")
110
如图所示,种群表现为多少是规律性的振荡,周期为6-7个世代。这个结果与无时滞的比较区别是如此之,表现在反馈作用上的一个世代时滞,就使稳定的种群增长型变成了有周期性的振荡或不稳定。分析其原因如下:由于时滞的影响,使密度对于增长率的影响在时间上后延了。因此,种群动态曲线的前部更凸了一些即增长更快了一些,随后,种群数量超过平衡密度还继续上升(因为时滞作用,使抑制性影响往后延),这称为超越(overshoot);然后种群数量骤然地下降,其下降也超过了平衡点,这称为超补偿(overcompen- sate),这样就形成了多少有周期性的振荡。
,这称为超越(overshoot);然后种群数量骤然地下降,其下降也超过了平衡点,这称为超补偿(overcompen- sate),这样就形成了多少有周期性的振荡。")
111
(二) 具有时滞的种群连续增长模型 连续增长的种群,最早提出具有时滞的逻辑斯谛增长方程的是美国学者赫琴森(Hutsinseg,1948),即: dN(t)/dt = r N t[(K – N(t - T))/K] 这个方程与前述逻辑斯谛增长方程不同之处在于(K – N(t - T))/K代替了(K – N)/K。在不具时滞的逻辑斯谛方程中,种群最大增长率的实现程度取决于t 时刻的N t值。而在具有时滞的逻辑斯谛方程中,它不取决于当时的种群密度N t 而是取决于更早时间的密度,即(t - T)时刻的种群密度N(t - T)。其中T称为反应时滞。 在逻辑斯谛方程中除了加进反应时滞,还可以引进生殖时滞,在一般水生生物中可以用怀卵期或其他同等的量来度量。
![(二) 具有时滞的种群连续增长模型 连续增长的种群,最早提出具有时滞的逻辑斯谛增长方程的是美国学者赫琴森(Hutsinseg,1948),即: dN(t)/dt = r N t[(K – N(t - T))/K]](http://slidesplayer.com/slide/11259827/61/images/111/%28%E4%BA%8C%29+%E5%85%B7%E6%9C%89%E6%97%B6%E6%BB%9E%E7%9A%84%E7%A7%8D%E7%BE%A4%E8%BF%9E%E7%BB%AD%E5%A2%9E%E9%95%BF%E6%A8%A1%E5%9E%8B+%E8%BF%9E%E7%BB%AD%E5%A2%9E%E9%95%BF%E7%9A%84%E7%A7%8D%E7%BE%A4%EF%BC%8C%E6%9C%80%E6%97%A9%E6%8F%90%E5%87%BA%E5%85%B7%E6%9C%89%E6%97%B6%E6%BB%9E%E7%9A%84%E9%80%BB%E8%BE%91%E6%96%AF%E8%B0%9B%E5%A2%9E%E9%95%BF%E6%96%B9%E7%A8%8B%E7%9A%84%E6%98%AF%E7%BE%8E%E5%9B%BD%E5%AD%A6%E8%80%85%E8%B5%AB%E7%90%B4%E6%A3%AE%EF%BC%88Hutsinseg%2C1948%EF%BC%89%EF%BC%8C%E5%8D%B3%EF%BC%9A+dN%EF%BC%88t%EF%BC%89%2Fdt+%3D+r+N+t%EF%BC%BB%28K+%E2%80%93+N%28t+-+T%29%29%2FK%EF%BC%BD.jpg "这个方程与前述逻辑斯谛增长方程不同之处在于(K – N(t - T))/K代替了(K – N)/K。在不具时滞的逻辑斯谛方程中,种群最大增长率的实现程度取决于t 时刻的N t值。而在具有时滞的逻辑斯谛方程中,它不取决于当时的种群密度N t 而是取决于更早时间的密度,即(t - T)时刻的种群密度N(t - T)。其中T称为反应时滞。 在逻辑斯谛方程中除了加进反应时滞,还可以引进生殖时滞,在一般水生生物中可以用怀卵期或其他同等的量来度量。")
112
这样模型可表示为: dN(t)/dt = r N t-g[(K – N(t - T))/K]
其中,g= 生殖时滞,T = 反应时滞。在其种群增长早期,生殖时滞对降低种群增长率有重要影响。 逻辑斯谛增长模型在引入时滞的概念后,具有了更为深刻的意义。生态学家应用计算机模拟技术获得的结果表明,逻辑斯谛方程在加进这种时滞以后,所产生的种群增长曲线有很多种变型。获得什么样的变型,取决于时滞的长短,但有一定的规律性。一般是在种群增长率r不变的情况下,时滞越长,系统的稳定性越低,即种群的数量变动越不稳定。
![这样模型可表示为: dN(t)/dt = r N t-g[(K – N(t - T))/K]](http://slidesplayer.com/slide/11259827/61/images/112/%E8%BF%99%E6%A0%B7%E6%A8%A1%E5%9E%8B%E5%8F%AF%E8%A1%A8%E7%A4%BA%E4%B8%BA%EF%BC%9A+dN%EF%BC%88t%EF%BC%89%2Fdt+%3D+r+N+t-g%EF%BC%BB%28K+%E2%80%93+N%28t+-+T%29%29%2FK%EF%BC%BD.jpg "其中,g= 生殖时滞,T = 反应时滞。在其种群增长早期,生殖时滞对降低种群增长率有重要影响。 逻辑斯谛增长模型在引入时滞的概念后,具有了更为深刻的意义。生态学家应用计算机模拟技术获得的结果表明,逻辑斯谛方程在加进这种时滞以后,所产生的种群增长曲线有很多种变型。获得什么样的变型,取决于时滞的长短,但有一定的规律性。一般是在种群增长率r不变的情况下,时滞越长,系统的稳定性越低,即种群的数量变动越不稳定。")
114
确切地表示为: 1. 在时滞值比较小时:0< T <e-1/r, 种群增长表现为逐渐地均匀地趋向于最大容纳量K值。
2.当时滞值增大时:e-1/r < T <л/2r, 种群会出现一定的波动,但波动的幅度越来越小,并趋于向稳定平衡点。 3.当时滞增加到一定值时: T >л/2r,种群增长就表现为具有周期性的数量波动,可以称为周期性振荡。 此外,某些时滞值也能产生减幅振荡,这种振荡是不稳定的,并可能导致种群的灭亡。 显然,这些结局与我们在自然界中所见到的种群动态更相吻合,从而使模型也就更加现实了。 具时滞的逻辑斯谛增长的数学模型能产生周期性数量波动,这个结果得到了许多实验种群研究的证实。

115
§6 种群数量变动及其生态对策 一、种群的数量变动 (一)种群数量的日变化 (二)、种群数量的季节消长 (三)种群数量的逐年变化
§6 种群数量变动及其生态对策 一、种群的数量变动 (一)种群数量的日变化 (二)、种群数量的季节消长 (三)种群数量的逐年变化 (四)种群数量的非周期性波动 二、种群的生态对策:r -对策者和K-对策者
种群数量的日变化. (二)、种群数量的季节消长. (三)种群数量的逐年变化. (四)种群数量的非周期性波动. 二、种群的生态对策:r -对策者和K-对策者.")
116
一、种群的数量变动 任何一种生物种群,都有不同程度的数量和生物量的变动。环境的变化越剧烈,种群数量的变化幅度也越大。当生活条件优良时,有机体不仅个体数量增加,而且生长加快,因而种群生物量也急剧增大;当生活条件恶化时,数量和生物量则降低,也就是说种群的再生产能力缩减到与具体条件相适应的水平。但是种群数量变化不是环境因素单方面作用的结果。各种生物都形成按照环境变化而调节的内源性反馈机制,数量变动是环境因素和种群适应性相互作用的结果。

117
(一)种群数量的日变化 生命周期短的种类,如细菌、单细胞藻类和原生动物,其繁殖和死亡常具有昼夜节律。如浮游生物昼夜垂直移动的同时,浮游生物种群数量的变化,扁平膝口藻,5:00数量开始减少,17:00降到最低点,以后又逐渐回升到半夜1:00达到最高值,可能与鲢鳙鱼的滤食节律有关。
种群数量的日变化 生命周期短的种类,如细菌、单细胞藻类和原生动物,其繁殖和死亡常具有昼夜节律。如浮游生物昼夜垂直移动的同时,浮游生物种群数量的变化,扁平膝口藻,5:00数量开始减少,17:00降到最低点,以后又逐渐回升到半夜1:00达到最高值,可能与鲢鳙鱼的滤食节律有关。")
118
(二)、种群数量的季节消长 在自然界中种群的变动,首先应区分出季节消长和年变动。种群的季节消长是人们所熟知的。各种生物所特有的种群季节消长特点,主要是受环境的季节性变化和生活史的适应性变化所调节。 由于太阳辐射的季节变化和随之而来的水温及水化学条件的变化,引起水体中生物种群数量的一系列变化。前已叙及,太阳照度的周期变化首先导致藻类种群的数量变化。藻类的数量变化又引起以藻类为食的植食动物种群的数量变化,以后肉食动物的数量也起了变化,因而产生了整个水体生物学的季节更替。种群数量和生物量的这种季节变化在极地和温带水体表现的最为显著。

119
1、浮游植物种群的季节变化 (1)冬季:冬季在低温、低光照条件下,浮游植物数量和产量都较低,但是在养分充足时,适低温和低光照的种类也能大量繁殖。但总之,由于水温低,繁殖减慢,以及冰下水层无涡流,而藻类细胞易下沉等原因,冬季浮游植物种群增长速度较慢。当冰层极厚和冰上覆雪时,可能停止增长和数量下降。冬季冰下种群通常由隐藻、蓝隐藻、光甲藻、衣藻、小球藻、锥囊藻、棕鞭藻、黄群藻、裸藻、囊裸藻、小环藻和菱形藻等。主要是那些能运动的小型种类。 (2)春季:冰溶后,水对流和风混合,产生水层的垂直环流,营养物质从底层上升,加上光照和温度的增高,为浮游植物的大量繁殖创造了良好条件。常出现硅藻种群的高峰,通常由1-2个优势种组成,常为直链藻、小环藻、冠盘藻、星杆藻等。一般高峰不超过3个月。
冬季:冬季在低温、低光照条件下,浮游植物数量和产量都较低,但是在养分充足时,适低温和低光照的种类也能大量繁殖。但总之,由于水温低,繁殖减慢,以及冰下水层无涡流,而藻类细胞易下沉等原因,冬季浮游植物种群增长速度较慢。当冰层极厚和冰上覆雪时,可能停止增长和数量下降。冬季冰下种群通常由隐藻、蓝隐藻、光甲藻、衣藻、小球藻、锥囊藻、棕鞭藻、黄群藻、裸藻、囊裸藻、小环藻和菱形藻等。主要是那些能运动的小型种类。 (2)春季:冰溶后,水对流和风混合,产生水层的垂直环流,营养物质从底层上升,加上光照和温度的增高,为浮游植物的大量繁殖创造了良好条件。常出现硅藻种群的高峰,通常由1-2个优势种组成,常为直链藻、小环藻、冠盘藻、星杆藻等。一般高峰不超过3个月。")
120
(3)夏季:硅藻种群数量降低,占优势的是绿藻,特别是绿球藻目的种群(盘星藻、卵囊藻等)。有时飞燕角甲藻也占优势。硅藻水华引起的水化学条件的改变(如溶解有机质和含氮量的增高)以及初夏光照和温度(20-25℃)等条件都适合绿藻种群的增长,水中含氮量降得极低,因此能固氮的蓝藻在仲夏取代绿藻而急剧增长。蓝藻占优势还与高温(>25℃)强光照、长日照、高pH值及较少被浮游动物滤食有关。如夏季大量出现的枝角类善于利用小型的绿藻,而难以利用大型的蓝藻。 (4)秋季:光照、温度均降,蓝藻大量死亡,出现硅藻种群的秋季次高峰,但不及春季。
秋季:光照、温度均降,蓝藻大量死亡,出现硅藻种群的秋季次高峰,但不及春季。")
121
2 浮游动物种群的季节变化 浮游动物种群数量的季节变化首先取决于浮游藻类和细菌的数量,其次决定于温度,此外还和本身的生活史以及食浮游动物的鱼类及其它动物摄食强度的季节节律有关。通常滤食性浮游动物的数量高峰在浮游植物之后。 浮游动物种群数量达到高峰后就由于被强烈摄食或本身死亡而下降。摄食者对浮游动物数量的影响在养鱼水体极为明显。 在淡水浮游动物中,桡足类是全年出现的,冬季在冰下水层越冬,春季随着水温的升高和营养条件的好转在水中数量增长而成为优势种。但是桡足类的生命周期较长,繁殖较慢,因此夏季以后常为出生率很强的的枝角类所取代。枝角类大多以冬卵越冬,冬卵必须积累一定热能才开始孵化,必须到春季夏末才出现较多的成体。夏季水温高,食物充足,单性生殖速度很快,种群数量迅速达到高峰。秋季降温以后又形成冬卵,数量急减而至于绝迹。轮虫和原生动物生命周期很短,其种群数量往往随食物条件而消长。

122
3. 底栖动物发展的季节性 底栖动物种群的季节变化首先与水温有关,水温既决定着它们的生殖期又影响起营养条件。其次也和被摄食强度的季节性有重要联系。在池塘和湖泊,底栖动物的数量有时还因出现窒死现象而急减;在河流中,水流速度的剧烈变化会破坏土壤的稳定性,可使底栖动物大为减少。例如,伏尔加河在1957 –1958 年冬季和早春,底栖动物生物量平均50 g/m2, 而洪水之后,由于土壤被搅动和生物被水流冲走,生物量降到0.003 g/m2。 水生昆虫的大量羽化也可能引起底栖动物数量的剧烈变动。在池塘和湖泊中,昆虫幼虫常占底栖动物总生物量的一半以上,这些幼虫的成群羽化可使底栖动物量降到极低点。按照主要昆虫所特有的世代数, 内陆水体底栖动物量一年可能出现一次到三﹑四次的波动。摇蚊幼虫的羽化时间有时十分集中,以至几周内几乎全部从底栖动物中消失。

123
(三)种群数量的逐年变化 种群在不同年份之间的变动,有的具有规律性(指有周期性的);有的没有规律性。有关种群动态的研究工作表明,大多数种类的多年数量动态表现为不规则的波动,有周期性数量变动的种类毕竟是有限的(不包括季节消长问题)。 对于水生生物而言,由于太阳辐射量的周期变化而产生的气候和水位的多年变动以及盐度﹑氧气等其它因素的年变化,都可以导致水生生物种群数量在不同年份的显著变化。由于太阳活动性的变化而引起的数量年变化,最常表现为9-11年的周期。 鱼类幼鱼的肥育条件及成活率和水位有密切关系,因此幼鱼期的水位情况对成鱼种群的丰度和渔获量常有明显的影响。

124
但是在很多情况下种群数量的年变化与自然地理周期的节律性无关,而是因捕食动物和被捕食动物种群相互作用的周期变化所引起的。当饵料生物数量很多时,捕食动物种群因营养条件的好转而增长。捕食动物的数量增长因增大了摄食量又导致饵料生物种群的减退,随之又恶化了本身的营养条件而使种群数量重新减少。捕捞的作用对鱼类和其它经济动物的数量变动也能起类似的作用;当资源量丰富时因容易捕捞,捕捞量也高,因捕捞过度导致资源量和捕捞量的下降,又为资源的再恢复创造了条件。 生命周期短的种类,数量的年变化表现得最为明显。要经过一段较长的时间浮游幼体发育期的种类,其数量年变化较之直接发育或浮游期短的种类更为剧烈。因为前者具有生殖量极高而个体易死亡的特点,在生活条件优良年份因幼体成活率高其数量急增,反之则急减。

125
(四)种群数量的非周期性波动 水体情况的非周期变化也引起种群数量的非周期性变动。例如水利建筑﹑污水排放等等均可引起这种变化,水体的富营养化引起的水生生物的改组和数量变化也属此类。
种群数量的非周期性波动 水体情况的非周期变化也引起种群数量的非周期性变动。例如水利建筑﹑污水排放等等均可引起这种变化,水体的富营养化引起的水生生物的改组和数量变化也属此类。")
126
二、种群的生态对策:r -对策者和K-对策者
任何一种生物的种群,都有不同程度的数量和生物量的变动。各种生物都能按照环境的变化而调节种群的数量,数量变动是环境因素和种群适应性相互作用的结果。 任何生物种群都具有相对稳定性。表现在种群的变动具有一定的上限和下限,还可能有一个长时间的平均水平(时滞)。当种群密度离开平均水平上升或下降时,就有一种使种群密度重返原来水平的倾向。并且种群数量上升或下降越剧烈,这种相反的倾向也越显著(即反馈调节)。
。当种群密度离开平均水平上升或下降时,就有一种使种群密度重返原来水平的倾向。并且种群数量上升或下降越剧烈,这种相反的倾向也越显著(即反馈调节)。")
127
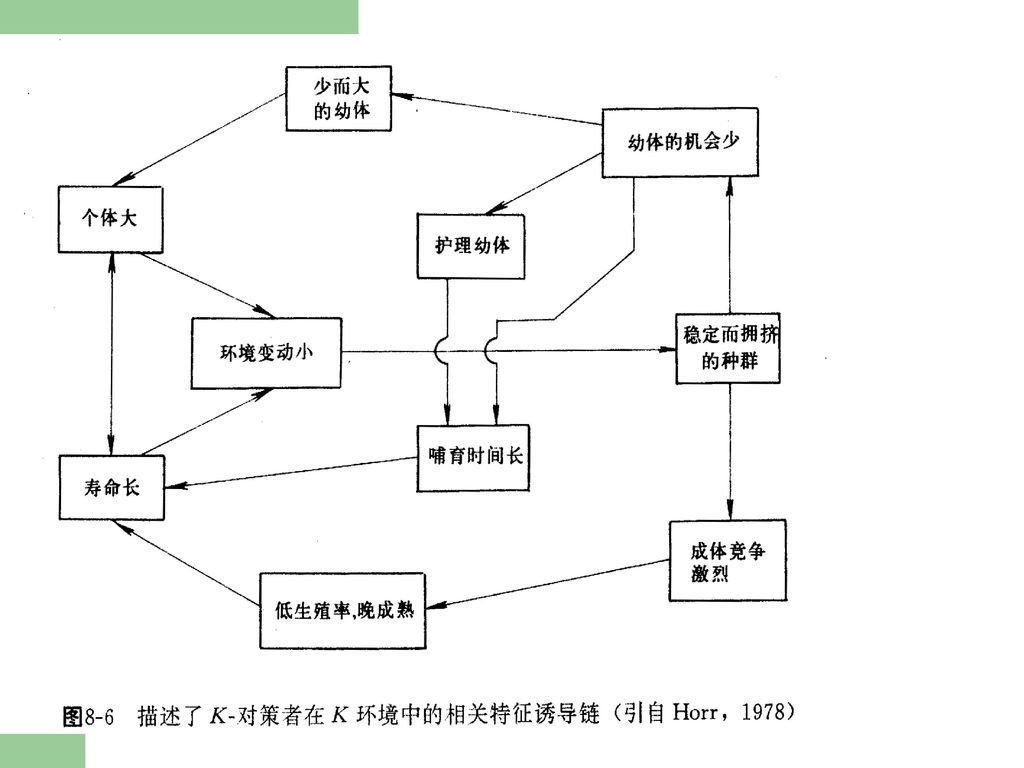
种群的变动和稳定性与物种的生态对策 即所谓的r-对策和K-对策有关。在复杂的自然环境条件下,各种生物都具有其独特的出生率﹑寿命﹑形体大小﹑存活率等生态特征。这些相互联系的生态特征,是物种在长期的进化过程中,与环境相互作用并通过自然选择而形成的。在生物进化过程中,按种群的动态类型形成了两类互不相同的适应。 一类称为r-选择(r-selected),这类生物它们的种群密度很不稳定,很少达到环境所能负载的最高负荷量——K值。一般保持在“S”型生长曲线的上升段。这类生物通常出生率高,寿命短,个体小,常常缺乏保护后代的机制,子代死亡率高,但具有较大的扩散能力。它们适应于不稳定的生活环境。另一类称为K-选择(K-selected),它们的种群密度比较稳定,经常处于K值周围,这类生物通常出生率低,寿命长,个体大,具有较完善的保持后代的机制,子代死亡率低,但一般扩散能力也较低。它们适应于稳定的生活环境。
,这类生物它们的种群密度很不稳定,很少达到环境所能负载的最高负荷量——K值。一般保持在 S 型生长曲线的上升段。这类生物通常出生率高,寿命短,个体小,常常缺乏保护后代的机制,子代死亡率高,但具有较大的扩散能力。它们适应于不稳定的生活环境。另一类称为K-选择(K-selected),它们的种群密度比较稳定,经常处于K值周围,这类生物通常出生率低,寿命长,个体大,具有较完善的保持后代的机制,子代死亡率低,但一般扩散能力也较低。它们适应于稳定的生活环境。")
128
从进化论观点来看,生物适应于不同栖息环境并朝着不同方向进化的“对策”称为生态对策(bionomic strategy),属于r-选择的生物可称为r-对策者(r-strategists);属于K-选择的生物可称为K-对策者(K-strategists)。r-对策者和K-对策者是两个进化方向不同的类型,在这中间还有一系列过渡的类型,有的更接近于r-对策者,有的更接近于K-对策者,也就是说从典型的r-对策者到典型的K-对策者之间有一个连续的谱系。 就生态分类单位而言,在水生生物中可以把鱼类和底栖动物视为K-选择的;而把浮游动物视为r-选择的。但是在每个大的生态类群中,也能有一个从r-选择到K-选择的谱系。例如在浮游动物中,原生动物﹑轮虫和枝角类均表现为r-对策者的特点,而桡足类则偏向于K-对策者;在鱼类中也有一些种类在r- K连续谱系中偏向r-选择的一端。

129
K-对策者的栖息环境是稳定的,在这种条件下增加种间斗争和竞争能力使种群密度保持在饱和密度附近是有利的,因此被选择的物种常常是个体大者。大个体由于增加世代周期而降低出生率,而出生率的降低必定要相应的增加存活率。因此这类动物防御和保护幼代的能力一般比较强。由于有亲代的抚育,出生率又低,也更容易达到长寿命和大的个体。K-对策者种群通常由许多不同的年龄世代组成,某一世代数量的变动可以从其它世代中得到平衡,但是由于r值较小,当种群遭受过度的死亡或激烈的动乱后,返回平衡水平的能力是比较低的。因此容易灭绝,对于这一类动物的资源应更加注意保护。 r-对策者的栖息环境是不稳定的﹑多变的,因而种群密度也难免处于经常激烈变动状态。个体小﹑寿命短有利于高的r值,能使它们在有利条件下迅速占领生境。因为它们占有的生境常是生态真空的,因而竞争能力不发达,对捕食者的防御能力比较弱,死亡率很高。但它们的高出生率可以弥补死亡的消耗。当种群密度过高和把资源耗尽时,这类动物具有较强的扩散能力,迁移恶化的生境到别处建立新的种群;有的种群则以休眠形式潜伏下来。

132
T.Southwood(1974)用增殖曲线说明了r/K两种对策者种群动态特征的区别:图中45°对角线表示子代Nt+1 与亲代Nt 相等,即种群处于平衡状态;对角线上方表示种群增长,Nt+1 > Nt;对角线下方表示种群减少,Nt+1 < Nt。K-对策种群增殖曲线呈“S”型,它与对角线有两个交点,一个是稳定平衡点S ,一个是不稳定平衡点X(又叫灭绝点),种群数量高于或低于平衡点S时,都趋于S(用两个收敛箭头表示),表示种群受到环境变动干扰后容易回到平衡点;但是在不稳定平衡点,两个发散箭头表示种群在此比较脆弱,容易因环境的变化而大起大落。与此相反,r-对策种群增殖曲线“J”型,它只有一个稳定平衡点S,而没有灭绝点;它们的种群在密度极低时,也能迅速回升到稳定平衡点S,并在S点上下波动。
用增殖曲线说明了r/K两种对策者种群动态特征的区别:图中45°对角线表示子代Nt+1 与亲代Nt 相等,即种群处于平衡状态;对角线上方表示种群增长,Nt+1 > Nt;对角线下方表示种群减少,Nt+1 < Nt。K-对策种群增殖曲线呈 S 型,它与对角线有两个交点,一个是稳定平衡点S ,一个是不稳定平衡点X(又叫灭绝点),种群数量高于或低于平衡点S时,都趋于S(用两个收敛箭头表示),表示种群受到环境变动干扰后容易回到平衡点;但是在不稳定平衡点,两个发散箭头表示种群在此比较脆弱,容易因环境的变化而大起大落。与此相反,r-对策种群增殖曲线 J 型,它只有一个稳定平衡点S,而没有灭绝点;它们的种群在密度极低时,也能迅速回升到稳定平衡点S,并在S点上下波动。")
133
由上述模式可见,r-对策种群与K-对策种群动态特征差异的本质在于rm。因为rm ≌ lnR0/T(其中R0 = N t+1/ Nt为净生殖率,T为平均世代时间),r-对策种群通过提高生殖率和缩短世代时间来达到高rm。在数量很低时,它们的R0 = N t+1/ Nt仍然 > 1,不象K-对策种群易于灭绝,而通过迅速增殖而恢复到较高水平,因此物种整体是富有恢复力的。K-对策种群则不同,当种群数量高于X时,种群尚能回升到S,一旦种群数量下降到某一临界密度(灭绝点)之下,种群生殖率R0 = N t+1/ Nt < 1,由此回到平衡水平的能力是有限的。如果确实很低,还有可能灭绝。这正是目前地球上珍稀动物所面临的状况。
,r-对策种群通过提高生殖率和缩短世代时间来达到高rm。在数量很低时,它们的R0 = N t+1/ Nt仍然 > 1,不象K-对策种群易于灭绝,而通过迅速增殖而恢复到较高水平,因此物种整体是富有恢复力的。K-对策种群则不同,当种群数量高于X时,种群尚能回升到S,一旦种群数量下降到某一临界密度(灭绝点)之下,种群生殖率R0 = N t+1/ Nt < 1,由此回到平衡水平的能力是有限的。如果确实很低,还有可能灭绝。这正是目前地球上珍稀动物所面临的状况。")
134
§7. 种群动态的调节机制 种群动态是反映出生和死亡、迁入和迁出这两组相互矛盾的过程,因此,所有影响出生、死亡和迁移的因素,都对种群动态有影响。

135
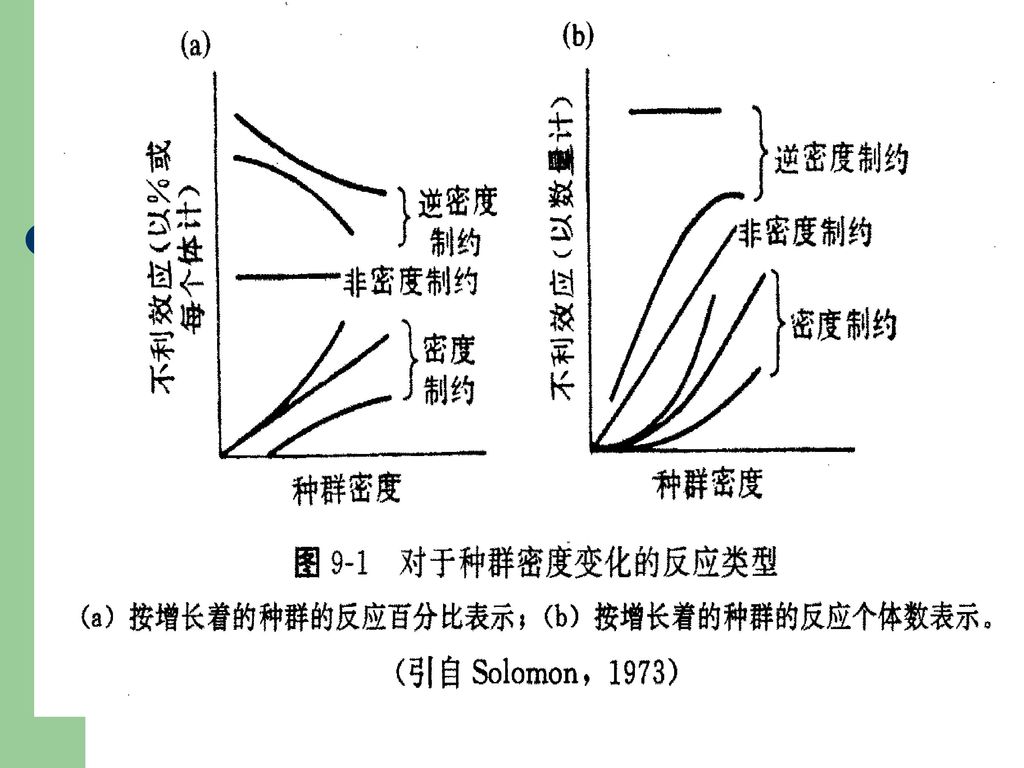
一、密度制约和非密度制约因素 霍华德和非斯克在研究引入寄生物来防治森林害虫时,将引起昆虫种群死亡的因素分为灾变性的(catastrophic)和兼性的(facultative),前者是不管种群密度如何,几乎总是杀死一定比例的个体,主要是气候因素;后者是指种群减少的比例是随着种群密度增加而增长的那些因素,所以霍华德和非斯克是种群调节的生物学派(biotic school)的雏形。通常把影响种群动态的因素粗略地区分为两大类史密斯(Smith, 1935):非密度制约因素和密度制约因素。 非密度制约(density-independent)因素,这类因素本身并不受种群密度所制约。即其作用的强度是独立于种群密度之外的。主要是一些非生物因素,如温度、盐度、气候等。反之,有利的气候条件,则可提高生物的出生率,增加种群的数量。但是,并不是所有的理化因素都不受种群密度的影响。
和兼性的(facultative),前者是不管种群密度如何,几乎总是杀死一定比例的个体,主要是气候因素;后者是指种群减少的比例是随着种群密度增加而增长的那些因素,所以霍华德和非斯克是种群调节的生物学派(biotic school)的雏形。通常把影响种群动态的因素粗略地区分为两大类史密斯(Smith, 1935):非密度制约因素和密度制约因素。 非密度制约(density-independent)因素,这类因素本身并不受种群密度所制约。即其作用的强度是独立于种群密度之外的。主要是一些非生物因素,如温度、盐度、气候等。反之,有利的气候条件,则可提高生物的出生率,增加种群的数量。但是,并不是所有的理化因素都不受种群密度的影响。")
136
密度制约(density—dependent)因素:
这类因素的作用强度随种群密度而变动,主要是生物性因素。包括种内关系和种间关系(捕食、竞争、寄生、共生、疾病等等)。 除上述划分的两类因素外,还把种群调节划分为内源性因素(内因)和外源性因素(外因)。前者指调节种群密度的原因在种群内部,即种内关系。外源性因素指调节种群密度的原因在于种群外部。 应当指出,决定种群动态的是因素的综合作用,而本是单个因素的作用。而强调因素的综合作用,并不否定某个单因素的主导作用,决定种群动态的主导因素可因情况不同而变化,也可以在一定条件下相对地不变。 因为种群动态的机制是极其复杂的,生态学家在这方面的讨论特别热烈,由此产生许多不同学说来解释种群动态的机理。
。 除上述划分的两类因素外,还把种群调节划分为内源性因素(内因)和外源性因素(外因)。前者指调节种群密度的原因在种群内部,即种内关系。外源性因素指调节种群密度的原因在于种群外部。 应当指出,决定种群动态的是因素的综合作用,而本是单个因素的作用。而强调因素的综合作用,并不否定某个单因素的主导作用,决定种群动态的主导因素可因情况不同而变化,也可以在一定条件下相对地不变。 因为种群动态的机制是极其复杂的,生态学家在这方面的讨论特别热烈,由此产生许多不同学说来解释种群动态的机理。")
138
二、非生物环境因素—气候学派 第一类决定种群动态的因素是理化环境因素,也是经常起作用、到处都存在的因素,它必然影响到生命活动,如生长、繁殖、死亡等各方面。理化环境条件(包括气候条件)决定着一种生物能否存活的大范围,而且在理化环境条件的变化中,影响着种群数量的波动。确有许多资料证明理化环境条件的改变与种群数量波动很好吻合。 强调非生物环境因家的即气候学派,最早提出气候是调节昆虫种群密度的是以色列的昆虫生物学家博登海默(F.B. Bidenheimer)。认为气候因子是种群数量变动的主要动因,反对自然种群处于稳定平衡的概念,强调野外种群的不稳定性。他们的观点是非密度制约论。 查普曼的生物潜能学说也可归之于气候学派。他认为生物种群有一固有的增殖能力,称为生物潜能或生殖潜能,但实际上不能实现,环境阻力降低了这种潜能,其公式:种群增长=生物潜能一环境阻力
决定着一种生物能否存活的大范围,而且在理化环境条件的变化中,影响着种群数量的波动。确有许多资料证明理化环境条件的改变与种群数量波动很好吻合。 强调非生物环境因家的即气候学派,最早提出气候是调节昆虫种群密度的是以色列的昆虫生物学家博登海默(F.B. Bidenheimer)。认为气候因子是种群数量变动的主要动因,反对自然种群处于稳定平衡的概念,强调野外种群的不稳定性。他们的观点是非密度制约论。 查普曼的生物潜能学说也可归之于气候学派。他认为生物种群有一固有的增殖能力,称为生物潜能或生殖潜能,但实际上不能实现,环境阻力降低了这种潜能,其公式:种群增长=生物潜能一环境阻力.")
139
三、生物因素——生物学派 生物学派主张密度制约因子如种间关系(捕食、寄生、竞争等过程对种群调节起决定作用。澳大利亚生物学家A.J. Nicholson。 食物因素也可以归为种间因素这一类,因为动物为异养生物,主要以其他动植物为食。 主张生物因子对种群调节起决定性作用的为生物学派,认为没有一个自然种群能无限制增长,因此,必然有许多限制种群增长的因素。一般情况下,种群也不致于下降到灭亡。因此,他们认为从长期来说,种群有一个平衡密度,即种群具有稳定性。

141
四、调和学派 生物学派认为生物因子(种间关系)是决定种群数量变动的主要因子,气候学派认为气候因子是主要因子。前者是密度制约论者,后者反对按密度制约性划分因素,认为一切因素都与密度有关。有的学者如A. Milne提出折衷的观点,主张气候因子和生物因子都具有决定种群密度的作用,当视具体条件而异。 由此,栖息于不稳定环境中的r—对策者更多地经受非密度制约因素的作用,栖息于有利的稳定的环境中的K一对策者则更多地经受密度制约因素的调节。

142
五、内源性因素-自我调节学派 内源性因素(自动调节因素)是指种群本身具有调节密度大小的各种因素,包括行为、生理和遗传等各种种内关系。 所有上面叙述的各种学说,都集中在外源性的因素上.都末涉及到种群自身的内部调节因素问题。他们的共同点就是把种群内所有的个体看作像物质的原子那样都是相同的,而忽视了种群内个体间的异质性。大量的研究表明,在种群内部存在“内在变化” ,这种变化在自然种群调节中可起重要作用。持这种观点的学派称为自我调节学派。按其论点不同又可分为行为调节学说,内分泌调节学说和遗传调节学说三派。

143
三派的共同点 ①自我调节学派强调节的内源性因素,而其他学派(包括气候学派、生物学派和调和学派)都是强调外因。强调外因的学派有一个共同特点,即把种群内的个体都看成是相同的,忽视种群内个体间的异质性。因此,自我调节学说在这个调节系统中,又加进一个自由度,即个体性质的变化。 ②自我调节学说是建立在种群内部的负反馈理论的基础上的,只有当密度增高,并超过平均密度时,种群自身的出生率降低.死亡率增高,或加强迁出等作用 即负反馈作用,才能使种群恢复或接近原有状态。反之,当密度向低于平均密度的方向偏离时,种群自身又通过加速生长发育,提高出生率,降低死亡率等反馈途径,使种群再恢复和接近原有水平。 ③种群自我调节学派的第三个共同特点是种群的自动调节。这是各物种所具有的适应性特征,这种适应性特征对于种内成员整体来说,能带来进化上的利益。因此,他将经受自然选择的考验。即种群数量增加时,通过自然选择压力和遗传组成的改变,为种群的减少,维持新的平衡铺平了道路。

144
(1)行为调节-温.爱德华学说 英国的V.C Wynne-Edwards提出行为调节学说:
社群行为就是一种调节种群的机制,如社群等级和领域性。 社群结构的建立是以种内个体间的支配一从属关系为基础。社群内有两种对立的倾向,一种是互相吸引,另一种是表现为社群中个体间的不相容,即反对过拥挤的一种倾向。这两种倾向都是结成社群结构的基础。而每一个动物在社群中占有一定的地位。领域性就是一个(或一对、一家)动物在一段时间中保卫其所占有的特定地盘,当种群密度超过一定限度时,领域的占领者就不允许同种其它个体入侵的行为。
动物在一段时间中保卫其所占有的特定地盘,当种群密度超过一定限度时,领域的占领者就不允许同种其它个体入侵的行为。")
145
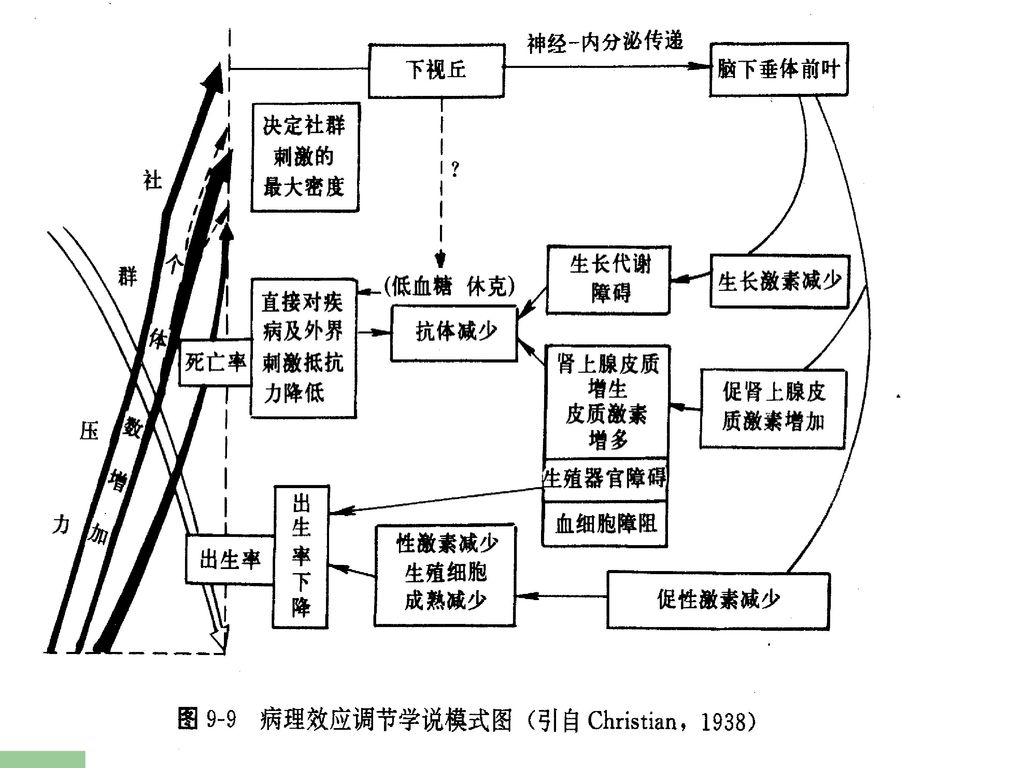
⑵内分泌调节-克里斯琴学说 JJ Christian认为当种群密度上升时,体内个体间的“紧张”(或称社群压力——social strees)增加,使内分泌系统产生变化,一方面使生长激素分泌减少.生长和代谢受到障碍。另一方面性激素分泌减少,生殖受到抑制,使出生率下降。它主要用来解释哺乳类的种群调节。
增加,使内分泌系统产生变化,一方面使生长激素分泌减少.生长和代谢受到障碍。另一方面性激素分泌减少,生殖受到抑制,使出生率下降。它主要用来解释哺乳类的种群调节。")
147
(3)遗传调节-奇蒂学说 D. Chitty认为种群内个体遗传型上有单的如遗传两型或遗传多态现象,其中一型在种群密度增加或密度高时占优势,另一型在下降时占伏势。当种群数量达到高峰时,由于社群压力增加,相互干扰加剧.自然选择有利于适应高密度但繁殖力低的一组基因型,种群数量就下降,这样进行遗传反馈,达到种群自我调节。遗传调节学说把生态学和遗传学联系起来,生态遗传学是目前许多学者注意的边缘科学。

148
结论 自动调节学派认为内源性因素是决定因素,将研究焦点放在种群内部各个个体成员之间的相互关系上。他们的共同前提是.种群密度影响种内的成员,使得出生率、死亡率和迁移发生改变,其共同观点是:种群调节是物种的一种适应性反应,这对于种群内成员整体来说,能带来进化上的利益,因此,它将经受自然的选择作用。

149
种群调节问题是最复杂的生态学理论问题。这是因为种群数量变动与许多内外因素有关、而且生物种类繁多、有的种群本身有较强的自我调节能力,有的这种调节能力较弱或根本没有调节能力,情况复杂。对这样复杂的问题,在初期工作阶段,各派学者在研究某个侧面并提出了自己的学说、片面性和局限性在所难免。由于许多学者的努力,逐渐由片面走向全面,由表面走向本质,由只强调外因而走向内因和外因相结合,这种总的发展趋势是十分明显的。

150
本节结束语 最后、我们用美国著名的生态学家Odum EP的一段话做为本节的结束语:“在结构简单,受物理压力或其它不规则或难以预计的外部影响较大的生态系统中,种群数量通常是受物理因素所调节、例如天气、水流、化学限制因子、污染等等。在结构复杂、不受物理胁迫的生态系统中,种群通常受生物因素所调节。所有生态系统,都表现出强烈的倾向,即通过自然选择,种群都向自我调节进化(因为种济数量过剩对任何神群都是不利的!),当然,这对于受外因控制的种群是较难达到的。”
,当然,这对于受外因控制的种群是较难达到的。")
151
思 考 题 1、何谓种群?说明它有哪些基本特征。 2、决定种群出生率的因素有哪些?试予以说明。 3、种群个体的空间分布有何特点?举例说明。
思 考 题 1、何谓种群?说明它有哪些基本特征。 2、决定种群出生率的因素有哪些?试予以说明。 3、种群个体的空间分布有何特点?举例说明。 4、什么是r-对策者和K-对策者?试比较两者种群动态特征的区别。 5、什么是内禀增长率?它与周限增长率有何关系?为什么? 6、什么是持续产量?利用Logistic增长理论如何确定最大持续产量? 7、种群数量变动的方式有哪几种形式?什么原因?

Similar presentations



 是在一定空间中同种个 体的组合的群。 这是最一般的定义,表示种群是由同种个体组 成的,占有一定的领域,是同种个体通过种内 关系组成的一个统一体或系统。除生态学外, 进化论、遗传学、分类学和生物地理学等都使 用种群这个术语。>")








.>")









