Download presentation
0
蛋白质降解及氨基酸代谢 Proteins Degradation& Amino acids Metabolism
宋潇达

1
一、甘油的氧化 脂肪动员产生的甘油主要在肝细胞经甘油激酶作用生成3-磷酸甘油,再脱氢生成磷酸二羟丙酮后循糖代谢途径分解或经糖异生途径转化成葡萄糖。脂肪细胞及骨骼肌等组织因甘油激酶活性很低,不能很好利用甘油。 二、脂肪酸的氧化分解 (一)脂肪酸的活化 在胞液中FFA通过与CoA酯化被激活,催化该反应的酶是脂酰CoA合成酶,需ATP、Mg2+参与。反应产生的PPi立即被焦磷酸酶水解,阻止了逆反应,所以1分子FFA的活化实际上消耗2个高能磷酸键。 RCOOH+ATP+CoASH—→RCO~SCoA+AMP+PPi (二)脂酰CoA进入线粒体 脂肪酸的氧化是在线粒体内进行的, 而脂酰CoA不能自由通过线粒体内膜进入基质, 需耍通过线粒体内膜上肉毒碱转运才能将脂酰基带入线粒体。内膜两侧的脂酰CoA肉毒碱酰基转移酶Ⅰ、Ⅱ(同工酶)催化完成脂酰基的转运和肉毒碱的释放。酶Ⅰ是FFA氧化分解的主要限速酶。 (三)脂酰CoA的β-氧化 脂酰CoA氧化生成乙酰CoA涉及四个基本反应:第一次氧化反应、水化反应、第二次氧化反应和硫解反应。
脂肪酸的活化. 在胞液中FFA通过与CoA酯化被激活,催化该反应的酶是脂酰CoA合成酶,需ATP、Mg2+参与。反应产生的PPi立即被焦磷酸酶水解,阻止了逆反应,所以1分子FFA的活化实际上消耗2个高能磷酸键。 RCOOH+ATP+CoASH—→RCO~SCoA+AMP+PPi. (二)脂酰CoA进入线粒体. 脂肪酸的氧化是在线粒体内进行的, 而脂酰CoA不能自由通过线粒体内膜进入基质, 需耍通过线粒体内膜上肉毒碱转运才能将脂酰基带入线粒体。内膜两侧的脂酰CoA肉毒碱酰基转移酶Ⅰ、Ⅱ(同工酶)催化完成脂酰基的转运和肉毒碱的释放。酶Ⅰ是FFA氧化分解的主要限速酶。 (三)脂酰CoA的β-氧化. 脂酰CoA氧化生成乙酰CoA涉及四个基本反应:第一次氧化反应、水化反应、第二次氧化反应和硫解反应。")
2
1. 奇数碳原子脂酸的氧化 人体含极少量奇数碳脂肪酸,而许多植物、海洋生物、石油酵母等含一定量的奇数碳脂肪酸。其β-氧化除生成乙酰CoA外,还生成1分子丙酰CoA,后者可通过β-羧化酶及异构酶的作用生成琥珀酰CoA,经TCA途径彻底氧化。 2. 不饱和脂肪酸的氧化 机体中脂酸约一半以上是不饱和脂肪酸,其中的双键均为顺式(cis)构型,不能被烯脂酰CoA水化酶作用(该酶催化的是反式构型双键的加水),所以需要异构酶和还原酶才能使一般不饱和脂肪酸的氧化进行下去。 三、酮体的生成和利用 脂肪酸经β-氧化生成的大多数乙酰CoA进入TCA循环,当乙酰CoA的量超过TCA循环氧化能力时,多余的生成酮体(ketone bodies),包括β-羟丁酸(占70%)、乙酰乙酸(占30%)和丙酮(微量)。 (一)酮体是在肝脏中合成的
构型,不能被烯脂酰CoA水化酶作用(该酶催化的是反式构型双键的加水),所以需要异构酶和还原酶才能使一般不饱和脂肪酸的氧化进行下去。 三、酮体的生成和利用. 脂肪酸经β-氧化生成的大多数乙酰CoA进入TCA循环,当乙酰CoA的量超过TCA循环氧化能力时,多余的生成酮体(ketone bodies),包括β-羟丁酸(占70%)、乙酰乙酸(占30%)和丙酮(微量)。 (一)酮体是在肝脏中合成的.")
3
2分子乙酰CoA经肝细胞线粒体乙酰乙酰CoA硫解酶催化缩合成乙酰乙酰CoA,再在羟甲基戊二酸单酰CoA合成酶(HMG- CoA合成酶)的催化下,结合第三个乙酰CoA生成β-羟基-β-甲基戊二酸单酰CoA。然后HMG- CoA裂解酶催化生成乙酰乙酸和乙酰CoA。(乙酰乙酰CoA也可在硫酯酶催化下水解为乙酰乙酸和CoA) (二)酮体的利用 酮体是正常的、有用的代谢物,是很多组织的重要能源。但肝细胞氧化酮体的酶活性很低,因此酮体经血液运输到肝外组织进一步氧化分解。心、肾、脑和骨胳肌线粒体有活性很高的氧化酮体的酶。β-羟丁酸在β-羟丁酸脱氢酶催化下重新脱氢生成乙酰乙酸,在不同肝外组织中乙酰乙酸可在琥珀酰CoA转硫酶或乙酰乙酸硫激酶作用下转变为乙酰乙酰CoA,再由乙酰乙酰CoA硫解酶裂解为2分子乙酰CoA,进入TCA途径彻底氧化。 脂肪的合成代谢 合成脂肪酸的酶系主要在胞浆,所以乙酰CoA需通过转运。合成脂肪酸的过程不同于β-氧化的逆过程,是由7种酶蛋白和酰基载体蛋白(ACP)组成的多酶复合体完成,合成的产物是软脂酸。碳链延长是在线粒体和内质网中的2个不同的酶系催化下进行的。
酮体的利用. 酮体是正常的、有用的代谢物,是很多组织的重要能源。但肝细胞氧化酮体的酶活性很低,因此酮体经血液运输到肝外组织进一步氧化分解。心、肾、脑和骨胳肌线粒体有活性很高的氧化酮体的酶。β-羟丁酸在β-羟丁酸脱氢酶催化下重新脱氢生成乙酰乙酸,在不同肝外组织中乙酰乙酸可在琥珀酰CoA转硫酶或乙酰乙酸硫激酶作用下转变为乙酰乙酰CoA,再由乙酰乙酰CoA硫解酶裂解为2分子乙酰CoA,进入TCA途径彻底氧化。 脂肪的合成代谢. 合成脂肪酸的酶系主要在胞浆,所以乙酰CoA需通过转运。合成脂肪酸的过程不同于β-氧化的逆过程,是由7种酶蛋白和酰基载体蛋白(ACP)组成的多酶复合体完成,合成的产物是软脂酸。碳链延长是在线粒体和内质网中的2个不同的酶系催化下进行的。")
4
(一)软脂酸的生物合成 乙酰CoA转运至胞浆(柠檬酸-丙酮酸循环)。 乙酰CoA羧化生成丙二酸单酰CoA 乙酰基和丙二酸单酰基的转移(负载) 脂肪酸合成酶系催化进行缩合、还原、脱水、还原反应。 哺乳动物脂肪酸氧化和合成的主要区别? (二)脂肪酸碳链的延长 (三)不饱和脂肪酸的合成 磷脂的生物合成 胆固醇的生物合成 1.乙酰CoA合成异戊烯焦磷酸(IPP) 2.鲨烯的合成 3.鲨烯转换为胆固醇 HMG- CoA还原酶(限速酶)
脂肪酸碳链的延长. (三)不饱和脂肪酸的合成. 磷脂的生物合成. 胆固醇的生物合成. 1.乙酰CoA合成异戊烯焦磷酸(IPP) 2.鲨烯的合成. 3.鲨烯转换为胆固醇. HMG- CoA还原酶(限速酶)")
5
目录 Section 1. 总览 Section 2. 蛋白质代谢 Section 3. 氨基酸分解 Section 4. 氨基酸合成
细胞外蛋白质降解 细胞内蛋白质降解 Section 3. 氨基酸分解 脱氨基反应 丙氨酸-葡萄糖循环 鸟氨酸循环 碳骨架代谢 个别氨基酸代谢 一碳单位 Section 4. 氨基酸合成 目录

6
第一部分. 总览 Nitrogen Cycle

7
氮的固定 固氮酶 Nitrogenase 对氮气的固定 谷氨酸脱氢酶 Glutamate dehydrogenase 通过碳骨架对氨的固定

8
The nitrogenase complex has two FeS enzymes:
氮的固定 The nitrogenase complex has two FeS enzymes: Nitrogenase reductase and Nitrogenase Reduces ~ 3 N2 /sec

9
Nitrogenase Reductase
Nitrogenase reductase is a dimer containing Fe4S4. It transfers electrons one at a time from ferredoxin to nitrogenase. This requires two ATP per e transferred.

10
Nitrogenase Nitrogenase is an a2b2 tetramer.
It commonly contains Mo, but some organisms use V. P cluster = Fe8S7 + 6 Cys Mo cluster = Fe7MoS9X + 1 Cys (X may be N.) a = red, b = blue
 a = red, b = blue.")
11
动物怎么办?

12
蛋白质,氨基酸代谢 NH4+ 细胞膜 葡萄糖 1946 诺贝尔奖 摄入的蛋白 ATP 氨基酸 三羧酸循环 氨基酸碳骨架 氨基酸库
蛋白酶消化 三羧酸循环 ATP 氨基酸 部分氨基酸的合成 氨基酸碳骨架 转运 溶酶体降解 降解 氨基酸库 门冬氨酸 1974,2013 诺贝尔奖 1953诺贝尔奖 富马酸 膜蛋白 胞外蛋白 胞内蛋白 泛素降解 合成 NH4+ 2004诺贝尔奖 2009诺贝尔奖 鸟氨酸循环 细胞内蛋白 尿素 蛋白质,氨基酸代谢 1953诺贝尔奖

13
第二部分. 蛋白质降解 细胞外蛋白质降解 蛋白质降解 细胞内蛋白质降解 不依赖于ATP的蛋白质降解 依赖于ATP的蛋白质降解 溶酶体介导
泛素介导 ⌘ 经验规则: N端规则(N- end rule);PEST序列 ⌘ 泛素途径: 蛋白酶体;高度调控
;PEST序列. ⌘ 泛素途径: 蛋白酶体;高度调控.")
14
(凝乳酶对乳有凝固作用,增加其在胃中停留时间,对婴儿很重要) 部位:小肠 PH:7 时间: 酶:
细胞外蛋白质降解(消化) 胃蛋白酶 多种酶 多肽、寡肽及少量氨基酸 氨基酸 蛋白质 部位:胃 PH:1-3 时间: 酶: 胃蛋白酶 Pepsin 胃分解蛋白酶 Gastricin 凝乳酶 chymosin (凝乳酶对乳有凝固作用,增加其在胃中停留时间,对婴儿很重要) 部位:小肠 PH:7 时间: 酶: 肽链外切酶:羧肽酶A、羧肽酶B、氨基肽酶、二肽酶 肽链内切酶:如胰蛋白酶Trypsin、糜蛋白酶Chymotrypsin、弹性蛋白酶 Elastase等。 100mL牛奶3mL凝乳酶溶液
 胃蛋白酶. 多种酶. 多肽、寡肽及少量氨基酸. 氨基酸. 蛋白质. 部位:胃. PH:1-3. 时间: 酶: 胃蛋白酶 Pepsin. 胃分解蛋白酶 Gastricin. 凝乳酶 chymosin. (凝乳酶对乳有凝固作用,增加其在胃中停留时间,对婴儿很重要) 部位:小肠. PH:7. 时间: 酶: 肽链外切酶:羧肽酶A、羧肽酶B、氨基肽酶、二肽酶. 肽链内切酶:如胰蛋白酶Trypsin、糜蛋白酶Chymotrypsin、弹性蛋白酶 Elastase等。 100mL牛奶3mL凝乳酶溶液.")
15
内切酶及外切酶 $ 内切酶 A 3 4 5 # A 6 K 7 2 8 Q 9 10 J $ # 2 3 5 4 6 A A 7 8 K 9
⌘ 内切酶:水解蛋白质肽链内部的一些肽键 ⌘ 外切酶:自肽链的末段开始每次水解一个氨基酸残基 扩展:实验中常用的内切酶(蛋白,核酸)
")
16
酶原 6 A K 7 7 K A A A 7 6 A A 10 K K 10 10 Q 10 Q 10 Q 10 Q J J J J
酶原,无活性 酶,有活性 胃酸,胃蛋白酶 胃蛋白酶 胃蛋白酶原 胰蛋白酶原 糜蛋白酶原 羧基肽酶原 弹性蛋白酶原 胰蛋白酶 糜蛋白酶 羧基肽酶 弹性蛋白酶 肠激酶 酶原:酶的前体 ⌘ 可保护组织免受蛋白酶的自身消化作用。 ⌘ 保证酶在其特定的部位和环境发挥催化作用。 ⌘ 酶原还可视为酶的贮存形式。

17
外源蛋白的吸收

18
必需氨基酸与非必需氨基酸 体内不能合成,必须由食物蛋白质供给的氨基酸称为必需氨基酸(essential amino acid)。 体内能够自行合成,不必由食物供给的氨基酸就称为非必需氨基酸。 必需氨基酸一共有八种:赖氨酸(Lys)、色氨酸(Trp)、苯丙氨酸(Phe)、蛋氨酸(Met)、苏氨酸(Thr)、亮氨酸(Leu)、异亮氨酸(Ile)、缬氨酸(Val)。 由于酪氨酸在体内需由苯丙氨酸为原料来合成,半胱氨酸必需以蛋氨酸为原料来合成,故这两种氨基酸被称为半必需氨基酸。 精氨酸、组氨酸在人体内合成慢,不能满足需要也称半必需氨基酸。
、色氨酸(Trp)、苯丙氨酸(Phe)、蛋氨酸(Met)、苏氨酸(Thr)、亮氨酸(Leu)、异亮氨酸(Ile)、缬氨酸(Val)。 由于酪氨酸在体内需由苯丙氨酸为原料来合成,半胱氨酸必需以蛋氨酸为原料来合成,故这两种氨基酸被称为半必需氨基酸。 精氨酸、组氨酸在人体内合成慢,不能满足需要也称半必需氨基酸。")
19
细胞内蛋白质(内源性)降解 ⌘ 蛋白在体内经常性被合成及分解. ⌘ 其半衰期从分钟到小时,到月。 ⌘ 与什么有关? 例子:
ornithine decarboxylase t½ ~11 min liver & plasma protein ~2-10 day muscle protein ~180 days collagen ~1000 days

20
细胞内蛋白质降解 N端规则(N-end rule) 蛋白质在细胞内的半衰期和它的N末端氨基酸种类有关
在80年代varshavsky alexander发现。 蛋白质在细胞内的半衰期和它的N末端氨基酸种类有关 Met, Ser, Ala, Thr, Val, 或 Gly 的蛋白,半衰期大于20小时 Phe, Leu, Asp, Lys, or Arg的半衰期小于3分钟

21
15种仅一个N端氨基酸不同的β- 半乳糖苷酶,在酵母中半衰期的差异从二十几个小时到2 min。
Alexander Varshavsky 科学发现的偶然性 15种仅一个N端氨基酸不同的β- 半乳糖苷酶,在酵母中半衰期的差异从二十几个小时到2 min。 N端氨基酸的种类似乎直接影响到这个蛋白在细胞内的稳定性。 检查了94个分泌性的毒蛋白,发现超过80%的蛋白,其N端氨基酸属于非常不稳定的一组,而一些其他分泌性蛋白也有这种倾向。 如果它们在制造过程中,不小心漏进细胞质内,就会很快地被分解掉。这个解释十分合理地说明细胞在制造分泌性毒蛋白的时候,为什么不会伤害到自己,因为毒蛋白在合成时,主要被局限在内质网里,在那里N端规则不适用,若有外泄则立刻被分解。

22
PEST序列(PEST sequence)
⌘ 蛋白富含Pro (P), Glu (E), Ser (S) and Thr (T), 叫做 PEST proteins, 在细胞内 更容易被降解,被认为是泛素化识别序列之一。
, Glu (E), Ser (S) and Thr (T), 叫做 PEST proteins, 在细胞内 更容易被降解,被认为是泛素化识别序列之一。")
23
溶酶体内降解过程 ⌘ 不依赖ATP ⌘ 利用组织蛋白酶(cathepsin)降解外源性蛋白、膜蛋白和长寿命的细胞内蛋白
降解外源性蛋白、膜蛋白和长寿命的细胞内蛋白")
24
自噬(autophagy)
")
25
泛素介导的降解过程(死亡标签) Ub 内吞作用, 溶酶体途径 Substrate 单泛素化 Ub Ub Ub E1 E2 Substrate
E3:Ring Ubiquitin Ligase Substrate 单泛素化 Ub Ub Ub E1 E2 Substrate Ub E3 Ub 多泛素化 Ub Ub E1:Ubiquitin Activating Enzyme E2:Ubiquitin Conjugating Enzyme Substrate 蛋白酶体降解 不降解 信号传导 细胞定位 逃脱死亡标签

26
泛素(Ubiqitin) ⌘ 76个氨基酸的小分子蛋白(8.5kD) ⌘ 普遍存在于真核生物细胞中 ⌘ 蛋白被泛素选择性标记后就会被降解
 ⌘ 76个氨基酸的小分子蛋白(8.5kD) ⌘ 普遍存在于真核生物细胞中 ⌘ 蛋白被泛素选择性标记后就会被降解")
27
泛素(Ubiqitin) ★ 泛素的C端羧基和目标蛋白Lys侧链e氨基形成异肽键 ★ 三个蛋白E1,E2,E3参与整个泛素化过程
 ★ 泛素的C端羧基和目标蛋白Lys侧链e氨基形成异肽键 ★ 三个蛋白E1,E2,E3参与整个泛素化过程")
28
泛素连接反应机理

29
反应机理 A.单泛素化: ATP ADP Ubiqutin specific protease

30
多泛素化链: C末端Gly与Lys48连接 碎纸机

31
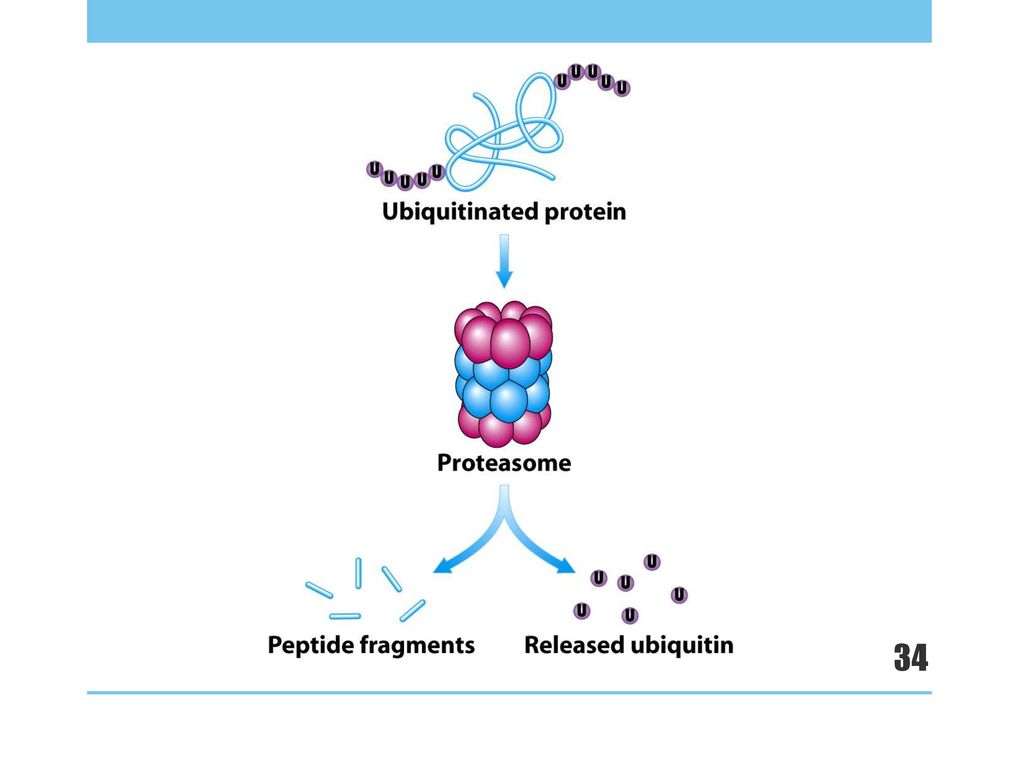
B.多泛素化: 同单泛素化 重复n次 Proteasome

32
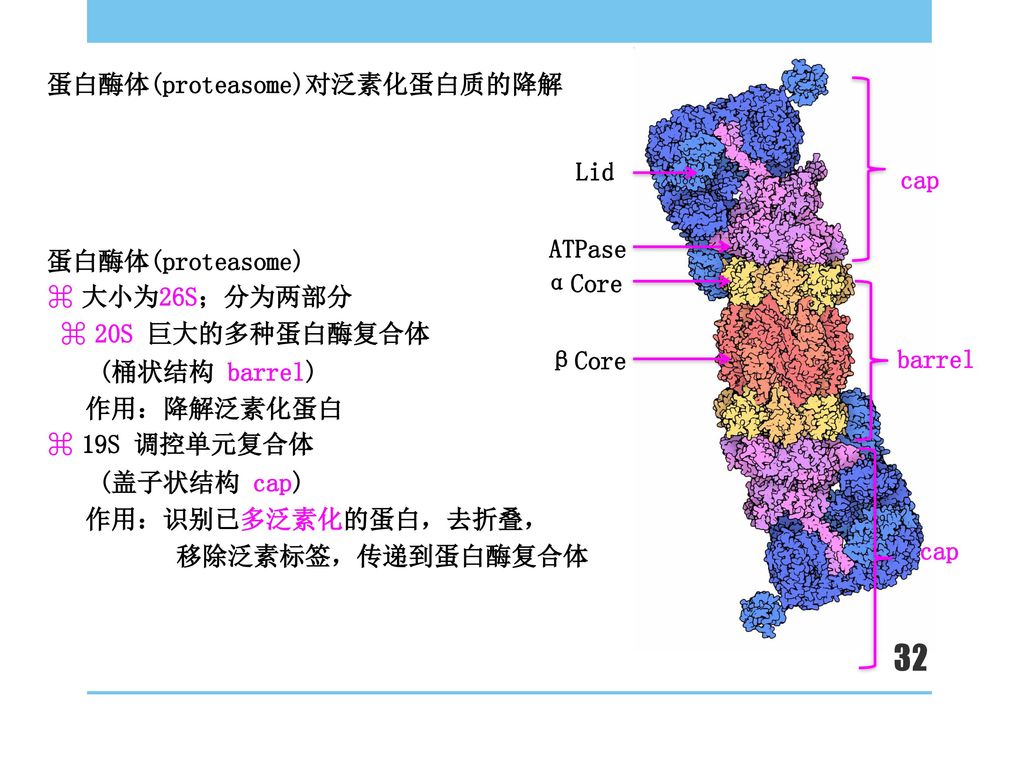
蛋白酶体(proteasome)对泛素化蛋白质的降解
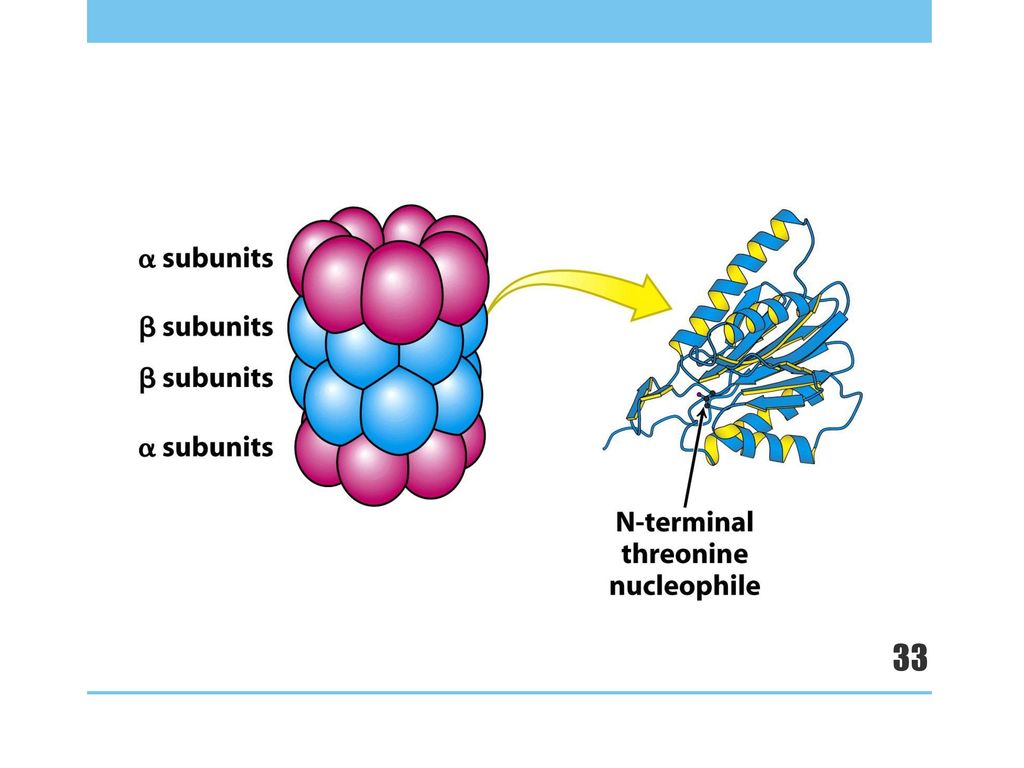
Lid cap ATPase 蛋白酶体(proteasome) ⌘ 大小为26S;分为两部分 ⌘ 20S 巨大的多种蛋白酶复合体 (桶状结构 barrel) 作用:降解泛素化蛋白 ⌘ 19S 调控单元复合体 (盖子状结构 cap) 作用:识别已多泛素化的蛋白,去折叠, 移除泛素标签,传递到蛋白酶复合体 αCore βCore barrel cap
 ⌘ 大小为26S;分为两部分. ⌘ 20S 巨大的多种蛋白酶复合体. (桶状结构 barrel) 作用:降解泛素化蛋白. ⌘ 19S 调控单元复合体. (盖子状结构 cap) 作用:识别已多泛素化的蛋白,去折叠, 移除泛素标签,传递到蛋白酶复合体. αCore. βCore. barrel. cap.")
36
Procaryotic vs Eucaryotic
Procaryotes have a proteasome analog of that in eucaryotes but the function is unclear since ubiquitin has not been found. In procaryotes, all α subunits are identical and all β subunits identical whereas in eucaryotes these subunits exhibit a number of isoforms. Procaryotes do have a ubiquitin-like protein but it is is used in the synthesis of thiamine and not protein degradation.

37
Procaryotic vs Eucaryotic

38
Procaryotic vs Eucaryotic
For thiamine synthesis For protein degradation

39
Section 3. 氨基酸分解 i FMNH2 骨架胺 少部分 骨架碳 α氨基酸 ii –H2O 二氧化碳 氨 iii 谷氨酸 ⌘ 氨基酸分解通常第一步为脱去氨基 ⌘ 碳原子骨架则进入能量代谢 ⌘ 氨基酸也用于新蛋白质的合成 ⌘ 长期饥饿的病人会降解氨基酸支持葡萄糖的异生

40
Section 3. 氨基酸分解 1. 氧化脱氨 (Oxidative deamination) Glu + NAD+ α-ketoglutarate + NADH + NH4+ 2. 转氨化 (Transamination) amino acid + α-ketoglutarate a-ketoacid + Glu 3. 直接脱氨 (Direct deamination) Ser pyruvate His urocanate (resonance driven) 4. 酰胺水解(Amide hydrolase) Gln or Asn Glu or Asp + NH4+
 Ser pyruvate. His urocanate (resonance driven) 4. 酰胺水解(Amide hydrolase) Gln or Asn Glu or Asp + NH4+")
41
1. 氧化脱氨 (Oxidative deamination)
L-amino acid oxidase Keto Acids FMNH2 NH3 L-Amino Acids FMN H2O FMNH2 O2 FMN H2O2 L-氨基酸氧化酶: ⌘ 体内分布很少 ⌘ 含有黄素蛋白(flavoprotein) ⌘ 产生氨 ⌘ 产生过氧化氢中间体 D-氨基酸氧化酶: ⌘ 体内分布广泛
 ⌘ 产生氨. ⌘ 产生过氧化氢中间体. D-氨基酸氧化酶: ⌘ 体内分布广泛.")
42
例:L谷氨酸脱氢酶 三羧酸循环 葡萄糖 ATP ⌘ ADP, GDP激活反应向右进行 ⌘ ATP, GTP促进反应向左进行 ⌘ 谷氨酸脱氢酶 (Mr 330,000) 线粒体基质 ⌘ 唯一一个同时识别NAD+,NADP+的酶

43
西弗碱中间体

44
2. 转氨化 (Transamination) (Thr, Pro, Lys例外) 绝大多数α氨基酸 α酮戊二酸 NADH + NH4+ 氨基转移酶 谷氨酸脱氢酶 NAD+ + H2O α酮酸 谷氨酸 转氨化 氧化脱氨化 联合脱氨化

45
氨基转移酶: 氨基转移酶: ✓催化不同α氨基酸和不同酮酸之间的氨基转移 ✓都带有辅基:磷酸吡哆醛(pyridoxal phosphate PLP) ✓都形成西弗碱中间体 吡哆醇 VB6 磷酸吡哆醛 PLP 磷酸吡哆胺 PMP

46
氨基转移酶 氨基转移酶作用机理: Lys 氨基转移酶 Lys258 Lys258 Lys

47
Bond Cleavage

48
例:天冬氨酸氨基转移酶(GOT,AST)
α酮戊二酸 草酰乙酸 谷氨酸 天冬氨酸氨基转移酶: 典型的转氨酶 常与丙氨酸氨基转移酶(GPT,ALT)用作诊断指标 GPT:诊断肝功能的正常与否;血清GPT高,可能急性肝炎 GOT:心脏病变的诊断;血清GOT高,可能心肌梗塞
用作诊断指标. GPT:诊断肝功能的正常与否;血清GPT高,可能急性肝炎. GOT:心脏病变的诊断;血清GOT高,可能心肌梗塞.")
49
3. 直接脱氨 丝氨酸 2-氨基丙烯酸 丙酮酸 苏氨酸 2-氨基-2-丁烯酸 α酮丁酸 Ser,Thr不需要通过谷氨酸即可直接脱氨。

50
4. 酰胺水解(Amide hydrolase)
Asparaginase and Glutaminase asparaginase asparagine > aspartate + NH4+ glutamine > glutamate + NH4+ glutaminase

51
氨的去毒化排除 生成氨基酸 少量 铵 水生动物 NH4+ 尿素 大多数陆生动物 尿酸 鸟类和爬行动物 ⌘ 过量的氨会引起人体内谷氨酰胺浓度增高导致脑部水肿, 同时会引起神经递质GABA浓度降低,引起细胞脱水症状。 ⌘ 氨在体内存在的形式?

52
谷氨酰胺与谷氨酰胺合成酶 ⌘ 氨去毒化的关键酶之一,尤其在脑组织,肾脏组织中。 ⌘ 谷氨酸的合成消耗大量的酮戊二酸。(酮戊二酸由三羧酸循环提供)干扰三羧酸循环。 ⌘ 谷氨酰胺合成酶受到严格的调控。 ⌘ 谷氨酰胺的酰胺氮可以用来合成其他氨基酸,嘌呤,嘧啶,以及多种氨基糖类。

53
谷氨酰胺合成酶的调控 ⌘ 变构调节---积累反馈抑制: 八种特殊的反馈抑制剂,包括谷氨酰胺的代谢终产物(色氨酸,组氨酸, 葡萄糖胺-6磷酸,氨甲酰磷酸,CTP,AMP),或者氨基酸代谢的指示剂 (丙氨酸和甘氨酸)每种抑制剂都可以绑定在酶的不同部位,同一个酶上绑定越多, 其抑制效应越强。 ⌘ 共价修饰---腺苷酰化: 由蛋白PII调控 ATP和酮戊二酸激活,谷氨酰胺抑制
每种抑制剂都可以绑定在酶的不同部位,同一个酶上绑定越多, 其抑制效应越强。 ⌘ 共价修饰---腺苷酰化: 由蛋白PII调控. ATP和酮戊二酸激活,谷氨酰胺抑制.")
54
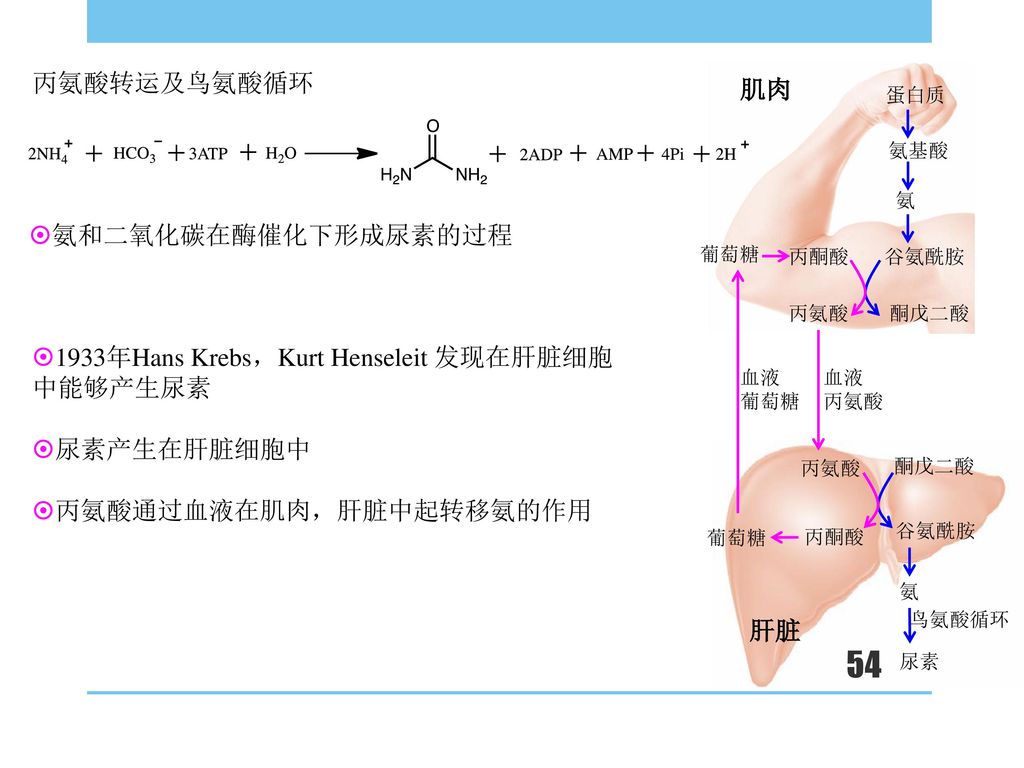
1933年Hans Krebs,Kurt Henseleit 发现在肝脏细胞中能够产生尿素
丙氨酸转运及鸟氨酸循环 肌肉 蛋白质 氨基酸 氨 氨和二氧化碳在酶催化下形成尿素的过程 葡萄糖 丙酮酸 谷氨酰胺 丙氨酸 酮戊二酸 1933年Hans Krebs,Kurt Henseleit 发现在肝脏细胞中能够产生尿素 尿素产生在肝脏细胞中 丙氨酸通过血液在肌肉,肝脏中起转移氨的作用 血液 葡萄糖 血液 丙氨酸 丙氨酸 酮戊二酸 丙酮酸 谷氨酰胺 葡萄糖 氨 鸟氨酸循环 肝脏 尿素

56
鸟氨酸循环的结构基础 氨甲酰磷酸 鸟氨酸 瓜氨酸 精氨酸
1932年,德国学者Krebs和Hensleit根据实验研究,提出了鸟氨酸循环(ornithine cycle)合成尿素的学说,这比三羧酸循环发现早5年。实验的根据是:将鼠肝切片置于胺盐和重碳酸盐介质中,有氧条件下保温数小时,发现胺盐含量减少,而尿素增多。当加入少量鸟氨酸、瓜氨酸或精氨酸能大大加速尿素的合成。肝脏又含有精氨酸酶,可催化精氨酸水解生成鸟氨酸和尿素。于是一个循环机制就出现。 瓜氨酸 精氨酸
合成尿素的学说,这比三羧酸循环发现早5年。实验的根据是:将鼠肝切片置于胺盐和重碳酸盐介质中,有氧条件下保温数小时,发现胺盐含量减少,而尿素增多。当加入少量鸟氨酸、瓜氨酸或精氨酸能大大加速尿素的合成。肝脏又含有精氨酸酶,可催化精氨酸水解生成鸟氨酸和尿素。于是一个循环机制就出现。 瓜氨酸. 精氨酸.")
57
鸟氨酸循环示意图 瓜氨酸 线粒体 细胞质 1 2 天冬氨酸 3 H2O 4 5 富马酸 瓜氨酸转运蛋白 鸟氨酸转运蛋白 ATP AMP
精氨基琥珀酸 精氨酸 鸟氨酸 细胞质 瓜氨酸转运蛋白 1 2 天冬氨酸 ATP 鸟氨酸转运蛋白 AMP 3 H2O 5 4 富马酸

58
鸟氨酸循环机理 鸟氨酸 瓜氨酸 精氨酸 富马酸 天冬氨酸 精氨基琥珀酸

59
Urea Cycle, Reaction 1 Transfer of the carbamoyl group in the mitochondria.

60
Urea Cycle, Reaction 2 A citrulline:ornithine antiport moves citrulline to the cytosol. The second NH2 for urea comes from Asp. An adenylated citrulline intermediate gives PPi.

61
Urea Cycle, Reaction 3 Cleavage of fumarate, production of arginine

62
Urea Cycle, Reaction 4 Cleavage of urea from arginine gives urea. The ornithine is ready to begin a new cycle.

63
Recycling to Aspartate
Malate dehydrogenase occurs in mitochondria and in the cytosol.

64
N-Acetylglutamate N-AcGlu is synthesized when NH4+ levels increase during amino acid catabolism. An activator of CPS I which provides substrate for the urea cycle. Glu is a product of Gln hydrolysis by CPS II in pyrimidine synth. An intermediate in ornithine synthesis.

65
Structural similarity
Blue - Ornithine transcarbamoylase from the urea cycle Red - Aspartate transcarbamoylase from pyrimidine synthesis

66
Functional similarity
Transfer of an amino group by incorporation of aspartate and elimination of fumarate occurs in both the urea cycle and pyrimidine synthesis.

67
鸟氨酸循环的调控 ⌘ 氨甲酰磷酸合成酶I 被N-乙酰谷氨酸变构激活 N-乙酰谷氨酸是由谷氨酸和乙酰辅酶A合成 ⌘ 其余酶是底物水平的调控 底物浓度增高激活鸟氨酸循环相关酶类

68
谢谢大家!


 不饱和脂肪酸的氧化 酮体 磷脂的代谢 脂肪酸代谢的调控.>")
 信号传导:激素、胞内信使.>")


 CO.>")




>")


蛋白质的降解: 外源蛋白的消化 内源性蛋白的选择性降解 (2)氨基酸的分解代谢:>")

物质在生物体的分解及合成代谢。要求学生重点掌握脂肪酸在生物体内的氧化分解途径—β-氧化和从头合成途径,了解脂类物质的功能和其他的氧化分解途径。 思考 脂类代谢 返回.>")




