Download presentation
1
Chap 脂类代谢 1 三酰甘油的分解代谢 2 三酰甘油的合成代谢 3 磷脂的代谢 4 胆固醇的代谢

2
脂类包括甘油三酯和类脂。甘油三酯是主要储能物质,类脂大都是细胞的重要结构物质和生理活性物质。 脂类的化学组成和结构差异很大,其共同特性是不溶于水而易溶于乙醚、氯仿等非极性溶剂的物质。通常脂类可按不同组成分为五类,即单纯脂、复合脂、萜类和类固醇及其衍生物、衍生脂类及结合脂类。

3
维生素A、D、E、K、固醇类激素(营养、代谢及调节)
脂类物质具有重要的生物功能。 脂肪是生物体中重要的贮藏物质。 1g蛋白质 kJ 1g糖 kJ 1g脂肪 kJ 脂类也是组成生物体的重要成分。 磷脂(生物膜)。 维生素A、D、E、K、固醇类激素(营养、代谢及调节) 机体表面的脂类(防止机械损伤与热量散发) 细胞表面的脂类(细胞识别、组织免疫) 脂类物质具有重要的生物功能。脂肪是生物体的能量提供者。1g脂肪彻底氧化可放出46.5kJ/mol能量,比1g糖或蛋白质放出的能量大一倍以上,因此脂肪是生物体内贮藏能量最多的物质。 每1g蛋白质、糖和脂肪所产生的代谢能量比较为: 1g蛋白质 kJ/mol 1g糖 kJ/mol 1g脂肪 kJ/mol 这样大的能量差异是因为脂肪是非极性的,它以近于无水的形式储藏着,而糖类和蛋白质均具有极性,它们以高度水合形式储藏着。1g干燥的糖元约结合2g水,所以实际上1g脂肪所贮存的能量为1g水合糖元贮存的六倍多。这就说明了为什么在进化过程中选择脂肪作为能量的储备形式。 脂类代谢主要讨论脂类在有机体内的降解和合成过程。了解脂类代谢对农业、工业、医学等方面都有重要的意义。例如种子的发芽率直接和种子的脂类代谢有关;又如利用微生物氧化石油中脂肪烃、工业生产低凝点油及其他化工产品;脂蛋白异常和威胁人类健康的冠心病等都与脂肪代谢关系密切。
。 维生素A、D、E、K、固醇类激素(营养、代谢及调节) 机体表面的脂类(防止机械损伤与热量散发) 细胞表面的脂类(细胞识别、组织免疫) 脂类物质具有重要的生物功能。脂肪是生物体的能量提供者。1g脂肪彻底氧化可放出46.5kJ/mol能量,比1g糖或蛋白质放出的能量大一倍以上,因此脂肪是生物体内贮藏能量最多的物质。 每1g蛋白质、糖和脂肪所产生的代谢能量比较为: 1g蛋白质 20.5kJ/mol. 1g糖 20.5kJ/mol. 1g脂肪 46.5kJ/mol. 这样大的能量差异是因为脂肪是非极性的,它以近于无水的形式储藏着,而糖类和蛋白质均具有极性,它们以高度水合形式储藏着。1g干燥的糖元约结合2g水,所以实际上1g脂肪所贮存的能量为1g水合糖元贮存的六倍多。这就说明了为什么在进化过程中选择脂肪作为能量的储备形式。 脂类代谢主要讨论脂类在有机体内的降解和合成过程。了解脂类代谢对农业、工业、医学等方面都有重要的意义。例如种子的发芽率直接和种子的脂类代谢有关;又如利用微生物氧化石油中脂肪烃、工业生产低凝点油及其他化工产品;脂蛋白异常和威胁人类健康的冠心病等都与脂肪代谢关系密切。")
4
1 三酰甘油的分解代谢

6
1.1 脂类的消化和吸收 脂肪的消化需要三种脂肪酶(脂肪酶、甘油二酯脂肪酶和甘油单酯脂肪酶 )参与,逐步水解甘油三酯的三个酯,最后生成甘油和脂肪酸。 脂肪降解的第一步是水解成甘油和脂肪酸,此反应由脂肪酶(简称脂酶,lipase)催化。组织中有三种脂肪酶:即脂肪酶、甘油二酯脂肪酶和甘油单酯脂肪酶,逐步把甘油三酯水解成甘油和脂肪酸。
催化。组织中有三种脂肪酶:即脂肪酶、甘油二酯脂肪酶和甘油单酯脂肪酶,逐步把甘油三酯水解成甘油和脂肪酸。")
7
脂类消化的主要场所是小肠。吸收后的脂类由脂蛋白参与转运代谢。食物中的脂肪在消化道内通过胆汁的乳化和脂肪酶的催化下分解成甘油和脂肪酸,经扩散的形式被小肠绒毛上皮细胞吸收后,在上皮细胞内再合成脂肪,与载脂蛋白结合后和类脂、胆固醇混合后一起进入毛细淋巴管,通过淋巴循环进入血液循环,最后输送到各部分组织。 脂类物质进人生物体内后的代谢去向:一是参与构成动物体的组织,如磷脂是生物膜的组成成分;二是作为备用物质贮存在皮下、肠系膜、大网膜等处;三是在需要的情况下,可再分解甘油、脂肪酸等,然后直接分解为CO2和H2O,或者转化成肝糖元等;四是被各种腺体利用来生成各自的特殊分泌物,如皮脂腺分泌的皮脂、乳腺所生成的乳汁等、内分泌腺分泌的各种类固醇激素等 在人和动物消化道内有脂肪酶,分解食物中的脂肪;以后甘油和脂肪酸在组织内再进一步氧化分解。植物也有类似的脂肪消化作用。如油料作物的种子萌发时,种子内脂肪酶活力增加,促使脂肪发生分解。凡能利用脂肪的微生物也都有脂肪酶,生产春雷霉素的培养基中需含有一定配比的植物油,说明春雷霉菌能够产生脂肪酶,所以能利用植物油,假丝酵母、圆酵母等都能产生较多的脂肪酶,工业上已经利用它们作为制造脂肪酶制剂的原料。 gallbladder 胆囊 bile salt:胆汁盐 emulsify:乳化 mucosa:胃粘膜 cholesterol :胆固醇 apolipoprotein :[生化]阿朴脂蛋白 chylomicron :[生化]乳糜微滴 lymphatic system :〈解〉淋巴系统 capillary: 毛细管;微血管

8
albumin 〈生化〉白蛋白,清蛋白 Serum:血清 hormone 荷尔蒙,激素 adipocyte【医】脂细胞,脂肪细胞

9
1.2 甘油代谢 甘油先与ATP作用,在甘油激酶(glycerol kinase)催化下生成α-磷酸甘油。然后再被氧化生成磷酸二羟丙酮,再经异构化,生成3-磷酸甘油醛,然后可经糖酵解途径转化成丙酮酸,进入三羧酸循环而彻底氧化,或经过糖异生途径合成糖元。因此甘油代谢和糖代谢的关系极为密切。
催化下生成α-磷酸甘油。然后再被氧化生成磷酸二羟丙酮,再经异构化,生成3-磷酸甘油醛,然后可经糖酵解途径转化成丙酮酸,进入三羧酸循环而彻底氧化,或经过糖异生途径合成糖元。因此甘油代谢和糖代谢的关系极为密切。")
11
脂肪酸的氧化 生物体内脂肪酸的氧化分解主要有-氧化、-氧化和-氧化等几条不同途径,-氧化途径最为重要和普遍。

12
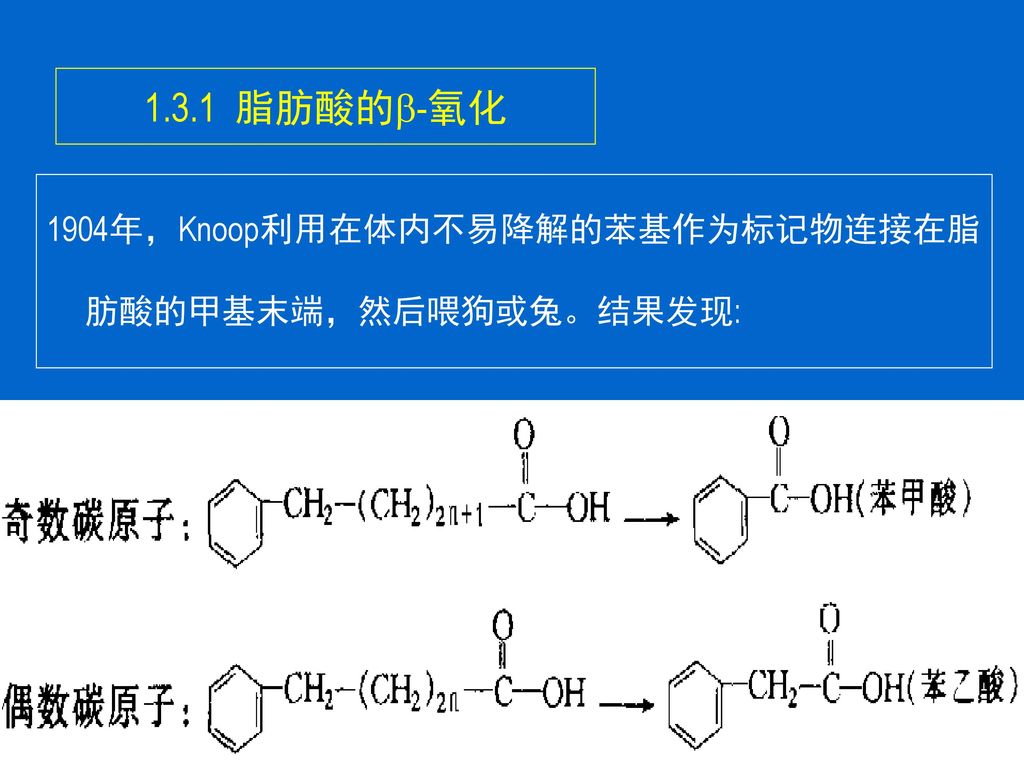
脂肪酸的-氧化 1904年,Knoop利用在体内不易降解的苯基作为标记物连接在脂肪酸的甲基末端,然后喂狗或兔。结果发现:

13
场所:线粒体基质(油料作物:乙醛酸循环体)。
由此提出脂肪酸的-氧化学说。 -氧化是从脂肪酸的羧基端-碳原子开始,碳链逐次断裂,每次产生1个乙酰CoA和比原来少2个C的脂肪酸链。 场所:线粒体基质(油料作物:乙醛酸循环体)。 -氧化是从脂肪酸的羧基端-碳原子开始,碳链逐次断裂,每次产生1个乙酰CoA和比原来少2个C的脂肪酸链。这是同位素示踪技术还未建立起来之前最具创造性的实验之一。
。 -氧化是从脂肪酸的羧基端-碳原子开始,碳链逐次断裂,每次产生1个乙酰CoA和比原来少2个C的脂肪酸链。这是同位素示踪技术还未建立起来之前最具创造性的实验之一。")
14
⑴脂肪酸的活化 活化形式:脂酰CoA。 酶:脂酰CoA合成酶 原料:CoA-SH、ATP, 脂肪酸。
脂肪酸在进行β-氧化降解前,在细胞质内必须先被激活成脂酰-CoA,该反应由脂酰-CoA合成酶(acyl-CoA synthetase)催化,需要ATP和CoA参与, 脂肪酸参加代谢前先要活化,活化形式是脂酰CoA。
催化,需要ATP和CoA参与, 脂肪酸参加代谢前先要活化,活化形式是脂酰CoA。")
15
脂酰CoA合成酶又称硫激酶,分布在细胞质、线粒体膜和内质网膜上。 细胞质内的硫激酶催化中短链脂肪酸活化;内质网膜上的酶活化长链脂肪酸,生成脂酰CoA,然后进入内质网用于甘油三酯合成;而线粒体膜上的酶活化的长链脂酰CoA,进入线粒体进入β-氧化。

16
⑵脂肪酸的转运 -氧化作用在线粒体基质进行,而在细胞质中形成的脂酰CoA不能透过线粒体内膜,需依靠内膜上的载体肉碱(即肉毒碱,3-羟-4-三甲氨基丁酸)携带,以脂酰基的形式跨越内膜而进入基质。
携带,以脂酰基的形式跨越内膜而进入基质。")
18
由于脂肪酸活化是在内质网或线粒体膜外,反应产物必须被转运至发生β-氧化作用的线粒体基质中,而脂酰-CoA不能直接穿过线粒体内膜,因此需要一个转运系统。转运脂酰-CoA的载体是肉毒碱(carnitine),即L-β羟基-γ-三甲基铵基丁酸,是一个由赖氨酸衍生而成的兼性化合物。它可将脂肪酸以酰基形式从线粒体膜外转运至膜内。其转运机制如下:肉毒碱与脂酰-CoA结合生成脂酰肉毒碱,该反应由肉毒碱脂酰转移酶(acyl-CoA transferase)催化,并在线粒体膜外侧进行,脂酰肉毒碱通过线粒体内膜的移位酶(translocase)穿过内膜,脂酰基与线粒体基质中的辅酶A结合,重新产生脂酰-CoA,释放肉毒碱。线粒体内膜内侧的肉毒碱转移酶 (acyl-CoA transferase)催化此反应。最后经肉毒碱移位酶协助,又回到线粒体外细胞质中(见图7-2)。
19
由于脂肪酸活化是在内质网或线粒体膜外,反应产物必须被转运至发生β-氧化作用的线粒体基质中,而脂酰-CoA不能直接穿过线粒体内膜,因此需要一个转运系统。转运脂酰-CoA的载体是肉毒碱(carnitine),即L-β羟基-γ-三甲基铵基丁酸,是一个由赖氨酸衍生而成的兼性化合物。它可将脂肪酸以酰基形式从线粒体膜外转运至膜内。其转运机制如下:肉毒碱与脂酰-CoA结合生成脂酰肉毒碱,该反应由肉毒碱脂酰转移酶(acyl-CoA transferase)催化,并在线粒体膜外侧进行,脂酰肉毒碱通过线粒体内膜的移位酶(translocase)穿过内膜,脂酰基与线粒体基质中的辅酶A结合,重新产生脂酰-CoA,释放肉毒碱。线粒体内膜内侧的肉毒碱转移酶 (acyl-CoA transferase)催化此反应。最后经肉毒碱移位酶协助,又回到线粒体外细胞质中(见图7-2)。
,即L-β羟基-γ-三甲基铵基丁酸,是一个由赖氨酸衍生而成的兼性化合物。它可将脂肪酸以酰基形式从线粒体膜外转运至膜内。其转运机制如下:肉毒碱与脂酰-CoA结合生成脂酰肉毒碱,该反应由肉毒碱脂酰转移酶(acyl-CoA transferase)催化,并在线粒体膜外侧进行,脂酰肉毒碱通过线粒体内膜的移位酶(translocase)穿过内膜,脂酰基与线粒体基质中的辅酶A结合,重新产生脂酰-CoA,释放肉毒碱。线粒体内膜内侧的肉毒碱转移酶 (acyl-CoA transferase)催化此反应。最后经肉毒碱移位酶协助,又回到线粒体外细胞质中(见图7-2)。")
20
酶Ⅰ使胞浆的脂酰CoA转化为辅酶A和脂酰肉毒碱,脂酰肉毒碱进入线粒体内膜。酶Ⅱ使脂肪酰肉毒碱转化成肉毒碱和脂酰CoA,肉毒碱重新发挥其载体功能,脂酰CoA则进入线粒体基质,成为脂肪酸β-氧化酶系的底物。

21
要吃就吃左旋肉碱吧 姐,我都饿得站不起来了 呵呵,咱姐三,就俺最有型

22
⑶ 氧化的生化历程 β氧化要经过脱氢、加水、再脱氢和硫解四步为一轮的循环反应,生成1分子乙酰CoA和一个少2个碳的脂酰CoA。

24
①脱氢反应 在脂酰CoA脱氢酶催化下,脂酰CoA在α和β碳原子上各脱去一个氢原子生成具有反式双键的α、β-烯脂酰辅酶A。

25
②加水反应 在烯酯酰CoA水化酶催化下,反式-2-烯脂酰-CoA在双键上加1分子水,生成β-羟脂酰CoA。

26
③脱氢反应 在β-羟脂酰CoA脱氢酶(辅酶为NAD)催化下,β-羟脂酰CoA脱氢生成β- 酮脂酰CoA。
催化下,β-羟脂酰CoA脱氢生成β- 酮脂酰CoA。")
27
④硫解反应 β-酮硫解酶催化在β-酮酯酰CoA的α、β碳原子之间断链,加上1分子辅酶A生成乙酰CoA和少2个碳原子的脂酰CoA。

28
β-氧化的4步反应都是可逆反应,但是第4步是高度放能的硫解反应,因此使整个β-氧化过程向裂解方向进行。 长链脂酰CoA经多次循环,每次降解下1分子乙酰CoA,直至成为2碳或3碳的脂酰CoA。 如软脂酸(棕榈酸,C15H31COOH)经7轮β-氧化生成8分子乙酰CoA。
经7轮β-氧化生成8分子乙酰CoA。")
30
⑷ 脂肪酸β-氧化的特点 ①脂肪酸活化生成脂酰CoA是耗能过程。中、短链脂肪酸直接进入线粒体,长链脂酰CoA需要肉毒碱转运。 ②β-氧化反应在线粒体内进行,没有线粒体的红细胞不能氧化脂肪酸供能。 ③β-氧化是需氧的过程,β-氧化过程中有FADH2和NADH生成,然后经呼吸链传递给氧,乙酰CoA的氧化也需要氧。

31
⑸脂肪酸β-氧化的能量变化 β-氧化是脂肪酸分解的主要途径,脂肪酸氧化可以供应机体所需要的大量能量,以18碳的饱和脂肪酸硬脂酸为例,其β-氧化的总反应为: CH3(CH2)15COSCoA + 8NAD+ + 8CoASH + 8H2O → 9CH3COSCoA + 8FADH2 + 8NADH + 8H+
15COSCoA + 8NAD+ + 8CoASH + 8H2O → 9CH3COSCoA + 8FADH2 + 8NADH + 8H+")
32
8分子FADH2提供8×2=16分子ATP; 8分子NADH+H+提供8×3=24分子ATP; 9分子乙酰CoA完全氧化提供9×12=108个分子ATP。 1 mol硬脂酸完全氧化生成CO2和H2O,共提供148 mol ATP。硬脂酸的活化过程消耗2 mol ATP,所以1 mol硬脂酸完全氧化可净生成146 mol ATP。

33
1 mol葡萄糖完全氧化可生成38分子ATP。3 mol葡萄糖所含碳原子数与1 mol硬脂酸相同, 3 mol葡萄糖可提供114mol ATP, 1 mol硬脂酸可提供146molATP。 在碳原子数相同的情况下脂肪酸能提供更多的能量。 脂肪酸氧化时释放出来的能量约有40%为机体利用合成高能化合物,其余60%以热的形式释出,热效率为40%。

34
脂肪酸β-氧化也是脂肪酸的改造过程,人体所需要的脂肪酸链的长短不同,通过β-氧化可将长链脂肪酸改造成长度适宜的脂肪酸,供机体代谢所需。 β-氧化过程中生成的乙酰CoA是重要的中间化合物,乙酰CoA除能进入三羧酸循环氧化供能外,还是许多重要化合物合成的原料,如酮体、胆固醇和类固醇化合物。

35
在油料种子萌发时,乙醛酸体中进行β-氧化产生的乙酰CoA一般不用于产能过程,而是通过乙醛酸循环转变为琥珀酸,再经糖异生作用转化为糖。

36
⑹不饱和脂肪酸的氧化 生物体内不饱和脂肪酸的双键都是顺式构型,第一个双键都在C9和C10之间,以后每隔3个碳原子出现1个。 不饱和脂肪酸的氧化与饱和脂肪酸基本相同,某些步骤还需异构酶的参与。

37
油酸(18:19)为单不饱和脂肪酸,它经历3轮-氧化作用后,产物在,位有一顺式双健,因此下一步反应不是脱氢,而是双键的异构化生成反式的,双键,然后-氧化作用继续正常进行。
为单不饱和脂肪酸,它经历3轮-氧化作用后,产物在,位有一顺式双健,因此下一步反应不是脱氢,而是双键的异构化生成反式的,双键,然后-氧化作用继续正常进行。")
39
因此油酸的氧化与相同碳的饱和脂肪酸(硬脂酸)相比,只是以一次双键异构化反应取代了一次脱氢反应,所以少产生1分子FADH2。 不饱和脂肪酸完全氧化生成CO2和H2O时提供的ATP少于相同碳原子数的饱和脂肪酸。 单不饱和脂肪酸和所有的多不饱和脂肪酸的前4轮-氧化作用都与油酸相同,都在第4轮时需要异构酶的参与。
相比,只是以一次双键异构化反应取代了一次脱氢反应,所以少产生1分子FADH2。 不饱和脂肪酸完全氧化生成CO2和H2O时提供的ATP少于相同碳原子数的饱和脂肪酸。 单不饱和脂肪酸和所有的多不饱和脂肪酸的前4轮-氧化作用都与油酸相同,都在第4轮时需要异构酶的参与。")
40
反烯酯酰辅酶A异构酶 顺二烯酯酰辅酶A还原酶

42
⑺奇数碳链脂肪酸的氧化 大多数脂肪酸含偶数碳原子,通过-氧化可全部转变成乙酰CoA,但一些植物和海洋生物能合成奇数碳脂肪酸,它们在最后一轮-氧化作用后,产生丙酰CoA。

43
先羧化,后经过两次异构化,形成琥珀酸。

44
丙酰CoA的代谢在动物体内代谢形成琥珀酰CoA进入三羧酸循环,继续进行代谢。 一些氨基酸,如异亮氨酸、缬氨酸和甲硫氨酸在降解过程中也会产生丙酰CoA或丙酸。 反刍动物瘤胃中的细菌可将糖类发酵产生大量的丙酸,而被宿主细胞吸收,丙酸先活化成丙酰CoA,然后进行代谢。

45
α-氧化 1956年,Stumpf,P.K. 在植物种子和叶子,动物的脑和肝细胞中发现了α-氧化作用。 在酶的催化下,在脂肪酸的-碳原子上发生氧化作用,分解出CO2,生成缩短了一个碳原子的脂肪酸,称为脂肪酸的-氧化。 Stumpf. P. K.(1956)发现植物线粒体中除有β-氧化作用外,还有1种特殊的氧化途径,称为α-氧化作用。这种特殊类型的氧化系统,首先发现于植物种子和植物叶子组织中,但后来也在脑和肝细胞中发现。在这个系统中,仅游离脂肪酸能作为底物,而且直接涉及到分子氧,每1次氧化经脂肪酸羧基端只失去1个碳原子,产物既可以是D-α-羟基脂肪酸,也可以是少1个碳原子的脂肪酸。
发现植物线粒体中除有β-氧化作用外,还有1种特殊的氧化途径,称为α-氧化作用。这种特殊类型的氧化系统,首先发现于植物种子和植物叶子组织中,但后来也在脑和肝细胞中发现。在这个系统中,仅游离脂肪酸能作为底物,而且直接涉及到分子氧,每1次氧化经脂肪酸羧基端只失去1个碳原子,产物既可以是D-α-羟基脂肪酸,也可以是少1个碳原子的脂肪酸。")
46
α-氧化是以游离脂肪酸作为底物,在-碳原子上发生羟化(-OH)或氢过氧化(-OOH),再进一步氧化脱羧。
脑苷脂和硫脂

47
长链脂肪酸在加单氧酶催化下生成α-羟脂肪酸,作为脑苷脂和硫脂的重要成分,α-羟脂肪酸继续氧化脱羧可生成奇数碳原子脂肪酸。 -氧化作用对于生物体内含甲基的支链脂肪酸的降解,或过长的脂肪酸(如C22、C24)的降解起着重要的作用。
的降解起着重要的作用。")
48
脂肪酸的-氧化途径 碳原子数在12个以上的脂肪酸可进行β-氧化,不产生二羧酸。但机体内也存在有少量的十二碳以下的脂肪酸,如十碳的癸酸和十一碳酸,通过ω-氧化途径进行氧化降解。 脂肪酸的ω-氧化作用:脂肪酸在混合功能氧化酶等酶的催化下,其ω碳(末端甲基碳)原子发生氧化,先生成ω-羟脂酸,继而氧化成α,ω-二羧酸的反应过程。 动物体内贮存的多是碳原子数在12个以上的脂肪酸,这些脂肪酸可进行β-氧化,不产生二羧酸。但机体内也存在有少量的十二碳以下的脂肪酸,如十碳的癸酸和十一碳酸,这些脂肪酸通过ω-氧化途径进行氧化降解。 脂肪酸的ω-氧化作用是指脂肪酸在混合功能氧化酶等酶的催化下,其ω碳(末端甲基碳)原子发生氧化,先生成ω-羟脂酸,继而氧化成α,ω-二羧酸的反应过程。ω omega欧米伽 在发现这一反应的初期,并未重视。目前ω-氧化酶系无论从理论上或实际上已日益受到重视,其原因是可利用它来清除海水表面的大量石油。反应过程是经浮油细菌的ω-氧化,把烃转变为脂肪酸,然后再进行脂肪酸两端的β-氧化降解。据估计,其氧化作用速率可高达0.5g/(d•m2),这对清除海面石油污染无疑会起重要作用。现已从油浸土壤中分离出许多细菌,它们具有ω-氧化酶系统,可用来清除海水表面的大量浮油。
原子发生氧化,先生成ω-羟脂酸,继而氧化成α,ω-二羧酸的反应过程。 动物体内贮存的多是碳原子数在12个以上的脂肪酸,这些脂肪酸可进行β-氧化,不产生二羧酸。但机体内也存在有少量的十二碳以下的脂肪酸,如十碳的癸酸和十一碳酸,这些脂肪酸通过ω-氧化途径进行氧化降解。 脂肪酸的ω-氧化作用是指脂肪酸在混合功能氧化酶等酶的催化下,其ω碳(末端甲基碳)原子发生氧化,先生成ω-羟脂酸,继而氧化成α,ω-二羧酸的反应过程。ω omega欧米伽. 在发现这一反应的初期,并未重视。目前ω-氧化酶系无论从理论上或实际上已日益受到重视,其原因是可利用它来清除海水表面的大量石油。反应过程是经浮油细菌的ω-氧化,把烃转变为脂肪酸,然后再进行脂肪酸两端的β-氧化降解。据估计,其氧化作用速率可高达0.5g/(d•m2),这对清除海面石油污染无疑会起重要作用。现已从油浸土壤中分离出许多细菌,它们具有ω-氧化酶系统,可用来清除海水表面的大量浮油。")
49
Succinate:琥珀酸 Adipate:己二酸

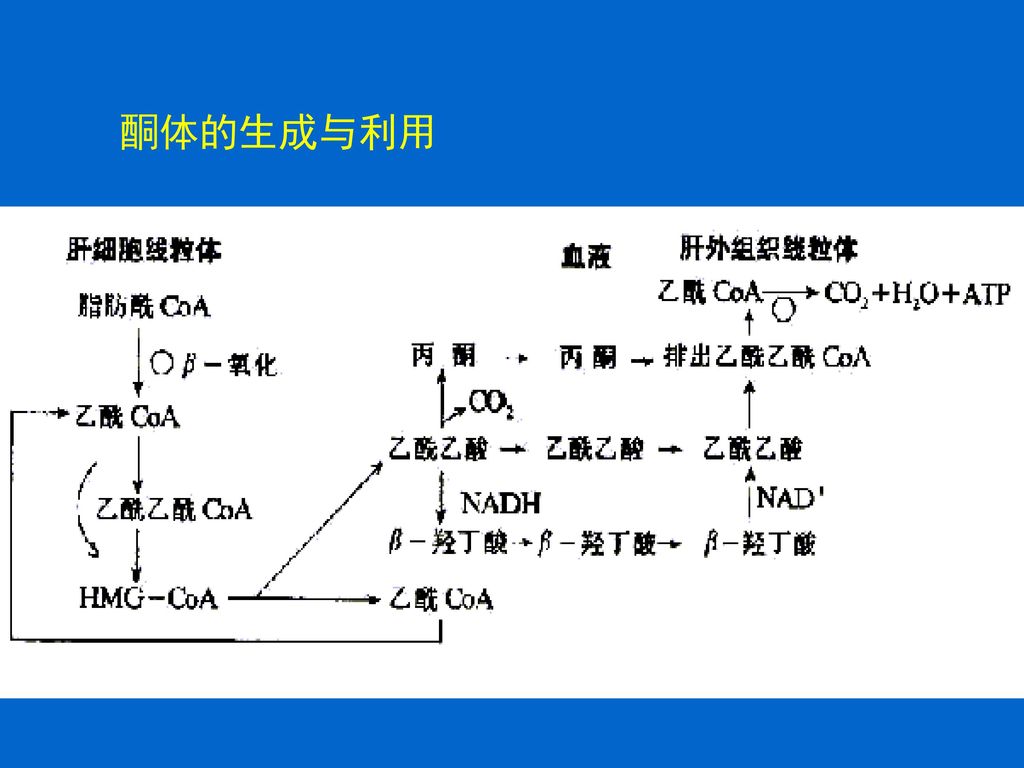
50
1.4 酮体代谢 脂肪酸-氧化及其它代谢所产生的乙酰CoA,在一般的细胞中可进入TCA进行氧化分解;而在肝脏中则可生成酮体。 酮体是脂肪酸在肝脏进行正常分解代谢所生成的特殊中间产物,包括乙酰乙酸(约占30%),β-羟丁酸(约占70%)和极少量的丙酮。
,β-羟丁酸(约占70%)和极少量的丙酮。")
52
酮体的合成 场所:酮体是在肝细胞线粒体中生成 原料:脂肪酸β-氧化生成的乙酰CoA。 酮体生成后迅速透过肝线粒体膜和细胞膜进入血液,转运至肝外组织利用。

56
酮体的分解 酮体在肝脏中产生后,必须由血液运送到肝外组织中进行分解。

57
酮体分解的关键是乙酰乙酸转变为乙酰乙酰CoA,反应由3-酮脂酰CoA转移酶催化,以琥珀酰CoA作为CoA的供体。 肝脏中缺乏3-酮脂酰CoA转移酶,因此只有在肝外组织中才能给乙酰乙酸加上CoA,然后裂解成乙酰CoA,从而通过TCA彻底氧化放能,也可作为合成脂肪酸的原料。

58
肝脏组织将乙酰CoA转变为酮体,而肝外组织则再将酮体转变为乙酰CoA。这并不是一种无效的循环,而是乙酰CoA在体内的运输方式。 肝脏组织正是以酮体的形式将乙酰CoA通过血液运送至外周器官中的。骨骼、心脏和肾上腺皮质细胞的能量消耗主要来自酮体,脑组织在糖饥饿时也能利用酮体作为能源。

59
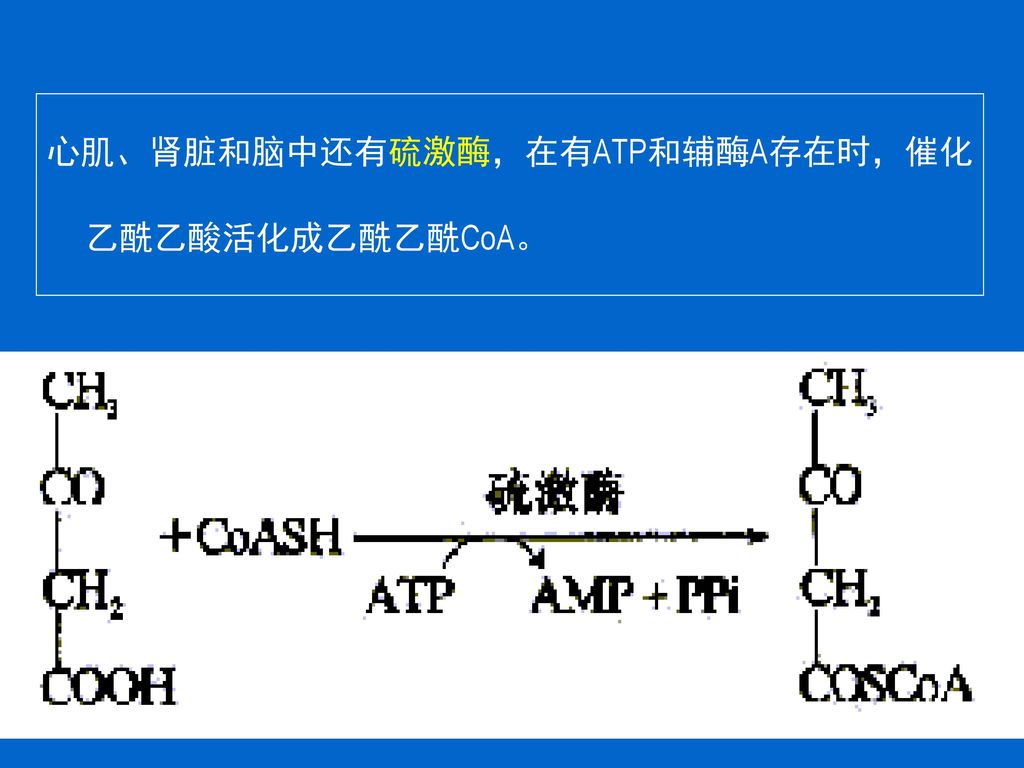
心肌、肾脏和脑中还有硫激酶,在有ATP和辅酶A存在时,催化乙酰乙酸活化成乙酰乙酰CoA。

60
乙酰乙酰CoA在硫解酶作用下,分解成两分子乙酰CoA,乙酰CoA主要进入TCA氧化分解。 丙酮除随尿排出外,有一部分直接从肺呼出,代谢上不占重要地位。

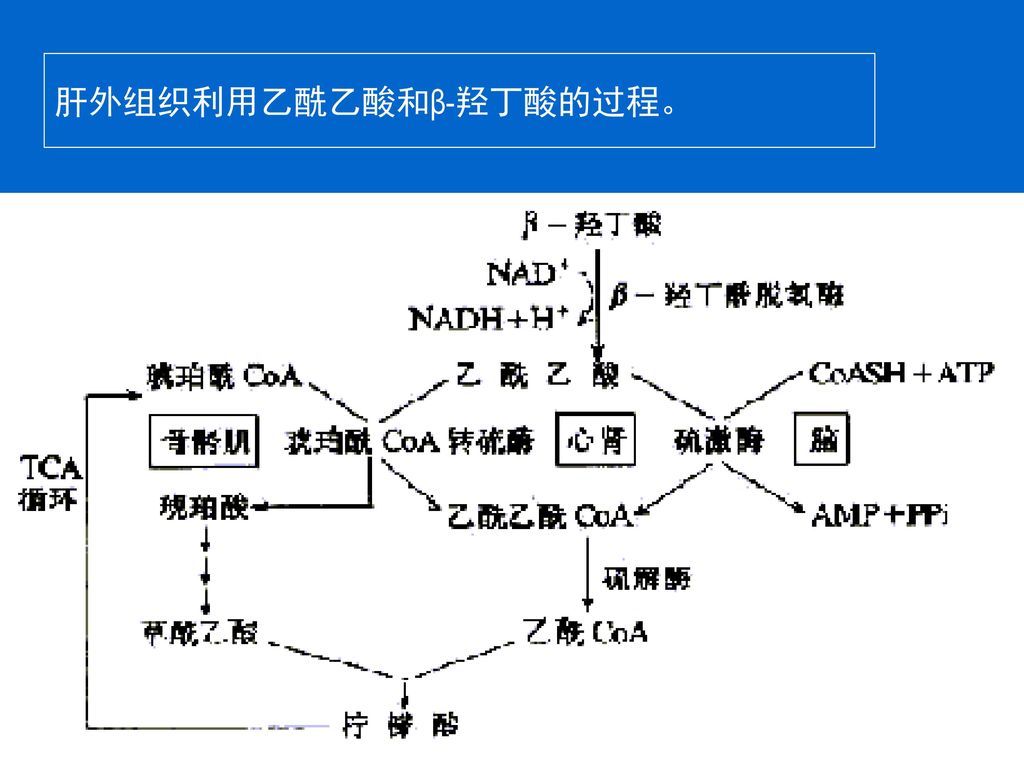
61
肝外组织利用乙酰乙酸和β-羟丁酸的过程。

62
1.4.3 酮体生成的意义 ①酮体易运输 酮体通过线粒体内膜以及在血中转运并不需要载体。 ②易利用 乙酰乙酸活化后只需一步反应就可以生成两分子乙酰CoA,β-羟丁酸的利用多一步氧化反应。 ③节省葡萄糖供脑和红细胞利用 肝外组织利用酮体氧化供能,减少了对葡萄糖的需求,同时乙酰CoA还能激活丙酮酸羧化酶,促进糖异生。

63
酮体的生成与利用

64
1.5 乙醛酸循环 乙醛酸循环存在于一些细菌、藻类和油料植物的种子的乙醛酸体中,它可将脂肪酸降解的主要产物乙酰CoA合成为琥珀酸。
1.5 乙醛酸循环 乙醛酸循环存在于一些细菌、藻类和油料植物的种子的乙醛酸体中,它可将脂肪酸降解的主要产物乙酰CoA合成为琥珀酸。 许多植物、微生物能够以乙酸为碳源合成其生长所需的其它含碳化合物,同时种子发芽时可以将脂肪转化成糖,这都是因为存在着一个类似于三羧酸循环的乙醛酸循环的缘故,该循环不存在于动物中。 乙醛酸循环从草酰乙酸和乙酰-CoA开始,形成柠檬酸后,异构化成异柠檬酸。与三羧酸循环不同的是异柠檬酸不经脱羧,而是被异柠檬酸裂解酶裂解成琥珀酸及乙醛酸。乙醛酸与另一个乙酰-CoA缩合形成苹果酸,此反应由苹果酸合酶催化,最后同三羧酸循环一样,苹果酸氧化成草酰乙酸,进入下一次循环。

65
乙醛酸循环的历程 ①乙酰CoA经柠檬酸合酶和顺乌头酸酶催化生成异柠檬酸 ②异柠檬酸裂解酶催化异柠檬酸裂解为乙醛酸和琥珀酸 ③在苹果酸合酶催化下乙醛酸和1分子乙酰CoA生成苹果酸 ④ 苹果酸经苹果酸脱氢酶催化生成草酰乙酸。整个过程构成一个循环反应。

67
总反应

68
乙醛酸循环可以看成是三羧酸循环的一个支路。 它在异柠檬酸处分支,绕过了三羧酸循环的两步脱羧反应,因而不发生氧化降解。参与乙醛酸循环的酶除了异柠檬酸裂解酶和苹果酸合酶外,其余的酶都与三羧酸循环的酶相同。 异柠檬酸裂解酶和苹果酸合酶是乙醛酸循环的关键酶。

69
乙醛酸循环的生物学意义 乙醛酸循环存在于一些细菌、藻类和油料植物的种子的乙醛酸体中。 油料植物的种子中主要的贮藏物质是脂肪,在种子萌发时乙醛酸体大量出现,由于它含有脂肪分解和乙醛酸循环的整套酶系,因此可以将脂肪分解,并将分解产物乙酰CoA转变为琥珀酸。

70
琥珀酸异生成糖并以蔗糖的形式运至种苗的其它组织,供给它们生长所需要的能源和碳源;而当种子萌发终止,贮脂耗尽,叶片能进行光合作用时,植物的能源和碳源可以由光和CO2获得,乙醛酸体的数量迅速下降以至完全消失。 对于一些细菌和藻类,乙醛酸循环使它们能够仅以乙酸盐作为能源和碳源生长。

71
植物线粒体内脂肪酸-氧化能力很低。乙醛酸循环将乙酰CoA转变为琥珀酸,再在线粒体中通过三羧酸循环的部分反应转化为苹果酸,然后进入细胞质,沿糖异生途径转变为糖类物质。

72
乙醛酸循环中也有苹果酸中间体,它也可以转运到细胞质中异生成糖,但需要及时回补,以保证循环的正常进行,苹果酸由TCA循环产物琥珀酸在线粒体中转变而来。 在脂肪转变为糖的过程中,乙醛酸循环起着关键的作用,它是连结糖代谢和脂代谢的枢纽。

73
2 脂肪的合成代谢 脂肪的生物合成包括甘油生成、脂肪酸生成以及甘油和脂肪酸分别转变为3-磷酸甘油和脂酰-CoA后的缩合三个步骤。

74
2.1 甘油的生物合成 甘油是由糖酵解的中间产物磷酸二羟丙酮在细胞质中合成。 与脂肪酸缩合成脂肪的是3-磷酸甘油,而不是游离的甘油。

75
脂肪酸的生物合成 脂肪酸多种多样,链的长短不一,不饱和键的数目、位置各不相同。 场所:细胞质 原料:乙酰CoA、ATP和NADPH 产物:先生成16C的软脂酸,经加工生成各种脂肪酸。 合成过程包括:①饱和脂肪酸的从头合成;②脂肪酸碳链的延长;③不饱和键的形成。 脂肪酸合成过程比较复杂,与氧化降解步骤完全不同。它可分为饱和脂肪酸的从头合成、脂肪酸碳链的延长和不饱和脂肪酸的生成几部分。脂肪酸合成的碳源主要来自糖酵解产生的乙酰-CoA,合成场所是在胞液中进行

76
2.2.1 饱和脂肪酸的从头合成 饱和脂肪酸的从头合成是在动物细胞质中进行的,植物则在叶绿体和前质体中进行。 过程:
饱和脂肪酸的从头合成 饱和脂肪酸的从头合成是在动物细胞质中进行的,植物则在叶绿体和前质体中进行。 过程: 乙酰CoA的穿梭(转运) 乙酰CoA的羧化(丙二酸单酰CoA的形成) 脂肪酸链的合成
 乙酰CoA的羧化(丙二酸单酰CoA的形成) 脂肪酸链的合成.")
77
⑴乙酰CoA的来源和转运 乙酰CoA是合成脂肪酸的主要原料,它来自丙酮酸氧化脱羧及氨基酸氧化等过程。这些代谢过程都是在线粒体内进行的,而脂肪酸合成发生在线粒体外。 乙酰CoA不能直接穿过线粒体内膜,需要在ATP供能,通过“柠檬酸穿梭”方式转移到线粒体外。 大部分脂肪酸合成定位于细胞质中,而脂肪酸β-氧化作用仅在线粒体中发生。脂肪酸合成所需碳源来自乙酰-CoA,但代谢产生的乙酰-CoA不能穿过线粒体的内膜到胞液中去,所以要借助“柠檬酸穿梭”来达到进入胞液的目的。柠檬酸穿梭途径是指乙酰-CoA与草酰乙酸结合形成柠檬酸,然后通过三羧酸载体透过膜,再由膜外柠檬酸裂解酶裂解成草酰乙酸和乙酰-CoA。草酰乙酸又被NADH还原成苹果酸再经氧化脱羧产生CO2 、NADPH和丙酮酸。丙酮酸进入线粒体后,在羧化酶催化下形成草酰乙酸,又可参加乙酰-CoA转运循环。

78
“柠檬酸穿梭”一次,使1分子乙酸CoA由线粒体进入胞液,同时消耗2分子ATP,还为机体提供了NADPH以补充合成反应的需要。

79
植物体中,线粒体内产生的乙酰CoA先脱去CoA以乙酸的形式运出线粒体,在线粒体外由脂酰CoA合成酶催化重新形成乙酰CoA。因此植物体内可能不存在“柠檬酸穿梭”过程

80
⑵ 丙二酸单酰CoA的生成 从头合成过程中,二碳单位的直接供体是丙二酸单酰CoA 。
在乙酰CoA羧化酶的催化下,消耗ATP,乙酰CoA和HCO3-反应形成丙二酸单酰CoA。

81
⑶脂肪酸合酶系统 脂肪酸合酶系统(FAS)是一个多酶复合体,它包括:①乙酰CoA:ACP转移酶、②丙二酸单酰CoA:ACP转移酶、③-酮脂酰-ACP合酶、④-酮脂酰-ACP还原酶、⑤-羟脂酰-ACP脱水酶、⑥烯脂酰-ACP还原酶等6种酶;还有脂酰基载体蛋白(ACP)作为辅助蛋白。 不同生物体内脂肪酸的合成过程相似,但FAS的组成却不相同。

83
不同生物体中的ACP十分相似,大肠杆菌中的ACP是一个由77个氨基酸残基组成的热稳定蛋白质,在它的第36位丝氨酸残基的侧链上,连有4-磷酸泛酰巯基乙胺。

84
-酮脂酰-ACP合酶 -酮脂酰-ACP还原酶 -羟脂酰-ACP脱水酶 丙二酸单酰CoA:ACP转移酶 乙酰CoA:ACP转移酶

85
除ACP上有一活性巯基外,-酮脂酰-ACP合酶上也有一活性巯基,由该酶多肽链上的一个半胱氨酸残基提供的,它是脂肪酸合成过程中另一个脂酰基载体。

86
因此,脂肪酸合酶系统上有两种活性巯基用于运载脂肪酸。
把ACP上的称中央巯基,-酮脂酰-ACP合酶上的称外围巯基。

87
⑷脂肪酸链的形成过程 脂肪酸链的形成过程是以乙酰CoA为起点,由丙二酸单酰CoA在羧基端逐步添加二碳单位,合成不超过16碳的脂酰基,最后脂酰基被水解成游离的脂肪酸。

88
整个过程都是在脂肪酸合酶系统中进行,由其中的酶和蛋白质协调完成。

89
缩合:在合成酶催化下外围巯基上的乙酰基与中央巯基上的丙二酸单酰基缩合成-酮丁酰基连接在中央巯基上,同时释放出一分子CO2。
①乙酰基和丙二酸单酰基进位(第一阶段) ②脂肪酸链延伸(第二阶段) 缩合:在合成酶催化下外围巯基上的乙酰基与中央巯基上的丙二酸单酰基缩合成-酮丁酰基连接在中央巯基上,同时释放出一分子CO2。 ①乙酰基和丙二酸单酰基进位(第一阶段) 乙酰基进位:乙酰CoA在转移酶催化下,乙酰基被转移到中央巯基上。 乙酰基移位:乙酰基由中央巯基转移到外围巯基上。 丙二酸单酰基进位:丙二酸单酰CoA在转移酶催化下,丙二酸单酰基被转移到中央巯基上。
 ②脂肪酸链延伸(第二阶段) 缩合:在合成酶催化下外围巯基上的乙酰基与中央巯基上的丙二酸单酰基缩合成-酮丁酰基连接在中央巯基上,同时释放出一分子CO2。 ①乙酰基和丙二酸单酰基进位(第一阶段) 乙酰基进位:乙酰CoA在转移酶催化下,乙酰基被转移到中央巯基上。 乙酰基移位:乙酰基由中央巯基转移到外围巯基上。 丙二酸单酰基进位:丙二酸单酰CoA在转移酶催化下,丙二酸单酰基被转移到中央巯基上。")
91
还原:在还原酶催化下,-酮丁酰基位羰基被NADPH还原成羟基,生成-羟脂酰基。

92
脱水:在脱水酶催化下-羟丁酰基的、碳原子间脱水生成反式2-烯丁酰基。

93
还原:在还原酶催化下2-烯丁酰基的双键被NADPH还原成单键生成延长了2个碳单位的丁酰基。

94
生成的丁酰基再与新进位的丙二酸单酰基重复缩合、还原、脱水、再还原的循环反应,又延长两个碳片段,生成己酯酰基,如此反复进行,直到生成软脂酰基为止。

96
③脂酰基水解(第三阶段) 当中央巯基上的脂酰基延长到一定程度(不超过16碳)后,在硫酯酶的作用下,ACP上的脂酰基或被转移到CoA上,或形成游离脂肪酸,或者直接用于合成磷脂酸。
 当中央巯基上的脂酰基延长到一定程度(不超过16碳)后,在硫酯酶的作用下,ACP上的脂酰基或被转移到CoA上,或形成游离脂肪酸,或者直接用于合成磷脂酸。")
97
由脂肪酸合酶系统形成1分子软脂酸需要消耗8分子乙酰CoA、7分子ATP和14分子NADPH。 NADPH主要来源于葡萄糖分解的磷酸戊糖途径。此外,苹果酸氧化脱羧也可产生少量NADPH。 (利用胞质中的乙酰CoA) 综上所述,脂肪酸合成每循环一次,碳链延长2个碳原子;CO2虽然在脂肪酸合成中参与起初的羧化反应,但在缩合反应中又重新释放出来,并没有消耗,它似乎仅仅起催化剂作用;在羧化反应中消耗ATP,此ATP由糖酵解提供;每次循环,经两次还原,消耗2NADPH+2H+,试验表明,脂肪酸合成需要的NADPH有60%是由磷酸戊糖途径提供的,其余部分可由糖酵解间接生成

99
⑸脂肪酸从头合成与-氧化的比较 脂肪酸的-氧化和从头合成过程不是简单的逆转关系。 它们反应的组织,细胞定位,转移载体,酰基载体,限速酶,激活剂,抑制剂,供氢体和受氢体以及反应底物与产物均不相同。

101
脂肪酸碳链的延长 脂肪酸的从头合成是在细胞质进行的,又称非线粒体系统合成途径,也称I型系统。因为β-酮脂酰ACP合成酶对软脂酰ACP无活性,所以由I型系统合成脂肪酸只能到生成16个碳的软脂酸为止。若要继续延长碳链,则需另外的延长系统途径,即线粒体(或微粒体)系统合成途径,称为Ⅱ型系统和Ⅲ型系统。
系统合成途径,称为Ⅱ型系统和Ⅲ型系统。")
102
延长过程发生在内质网以及动物的线粒体和植物的叶绿体或前质体中。不同的部位延长的具体方式都不相同。

103
在植物中,软脂酸的碳链延长在细胞质中进行,由延长酶系统Ⅱ和Ⅲ催化,形成18碳、20碳的脂肪酸
在植物中,软脂酸的碳链延长在细胞质中进行,由延长酶系统Ⅱ和Ⅲ催化,形成18碳、20碳的脂肪酸. 植物体内的脂肪酸合成酶系统至少有三个类型,其作用如下式:

104
动物:软脂酸碳链的延长在内质网(微粒体)或线粒体中进行。 内质网上的延长以软脂酰-CoA为基础。以丙二酸单酰-CoA为二碳供体,以CoA-SH为酰基载体,经过缩合、还原、脱水和再还原,生成硬脂酰-CoA。然后重复循环,生成20碳以上的脂酰-CoA。 线粒体中软脂酸的延长是与β-氧化相似的逆向过程:以软脂酰-CoA与乙酰-CoA (二碳供体)进行缩合、还原、脱水和再还原,生成硬脂酰-CoA。重复循环,可继续加长碳链(延长到C24至 C26)。
或线粒体中进行。 内质网上的延长以软脂酰-CoA为基础。以丙二酸单酰-CoA为二碳供体,以CoA-SH为酰基载体,经过缩合、还原、脱水和再还原,生成硬脂酰-CoA。然后重复循环,生成20碳以上的脂酰-CoA。 线粒体中软脂酸的延长是与β-氧化相似的逆向过程:以软脂酰-CoA与乙酰-CoA (二碳供体)进行缩合、还原、脱水和再还原,生成硬脂酰-CoA。重复循环,可继续加长碳链(延长到C24至 C26)。")
105
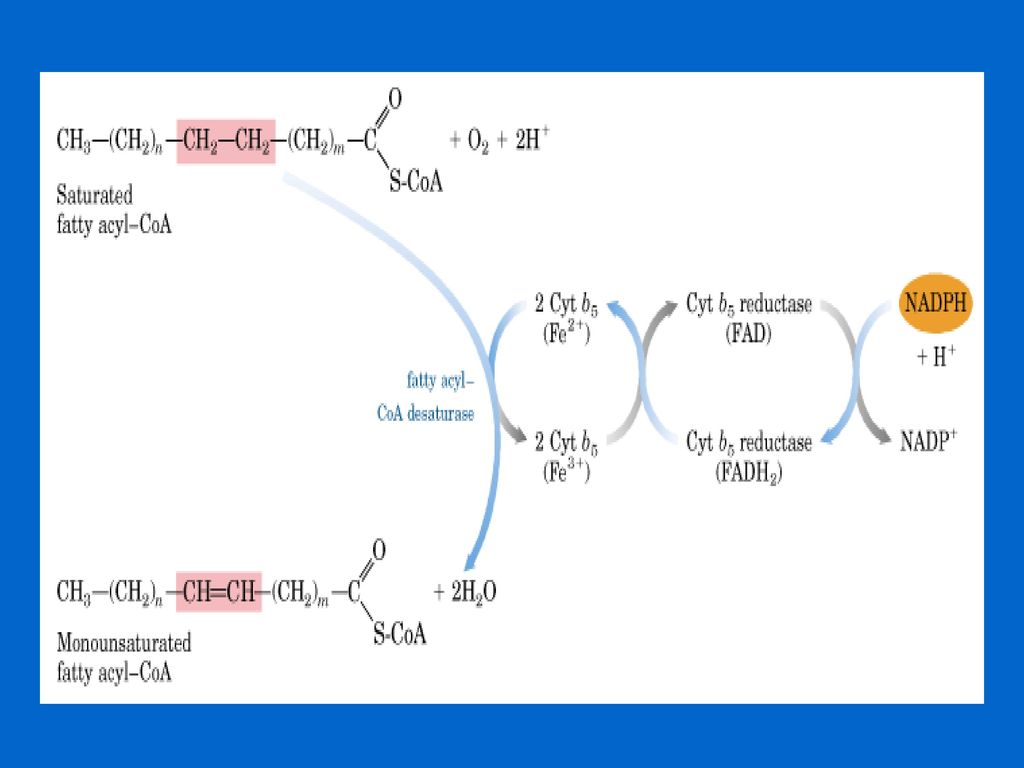
脂肪酸碳链的去饱和 生物体内存在大量的各种不饱和脂肪酸,如棕榈油酸(16:1 9)、油酸(18:l9)、亚油酸(18:29 ,12)、亚麻酸(18:39,12,15)等,它们都是由饱和脂肪酸经去饱和作用而形成的。 去饱和作用主要有需氧途径(真核生物)和厌氧途径(厌氧微生物) 哺乳动物主要有四类不饱和脂肪酸: 棕榈油酸(ω-7):16:1 Δ 十六碳单烯脂酸,双键位于第9位 油酸 (ω-9):18:1 Δ 十八碳单烯脂酸,双键位于第9位 亚油酸 (ω-6):18:2 Δ9,12 十八碳二烯酸, 双键位于第9,12位 亚麻酸 (ω-7):18:3 Δ9,12,15 十八碳三烯酸, 双键位于第9,12,15位 其中亚油酸和亚麻酸是人体必需脂肪酸,因为人和其他哺乳动物缺乏在脂肪酸第9位碳原子以上位置引入双键的酶系,所以自身不能合成亚油酸和亚麻酸,必须从植物中获得。亚油酸和亚麻酸广泛存在于植物油(花生、芝麻和棉籽油等)中。其他多不饱和脂肪酸都是由以上4种不饱和脂肪酸衍生而来,通过延长和去饱和作用交替进行来完成的。
、油酸(18:l9)、亚油酸(18:29 ,12)、亚麻酸(18:39,12,15)等,它们都是由饱和脂肪酸经去饱和作用而形成的。 去饱和作用主要有需氧途径(真核生物)和厌氧途径(厌氧微生物) 哺乳动物主要有四类不饱和脂肪酸: 棕榈油酸(ω-7):16:1 Δ9 十六碳单烯脂酸,双键位于第9位. 油酸 (ω-9):18:1 Δ9 十八碳单烯脂酸,双键位于第9位. 亚油酸 (ω-6):18:2 Δ9,12 十八碳二烯酸, 双键位于第9,12位. 亚麻酸 (ω-7):18:3 Δ9,12,15 十八碳三烯酸, 双键位于第9,12,15位. 其中亚油酸和亚麻酸是人体必需脂肪酸,因为人和其他哺乳动物缺乏在脂肪酸第9位碳原子以上位置引入双键的酶系,所以自身不能合成亚油酸和亚麻酸,必须从植物中获得。亚油酸和亚麻酸广泛存在于植物油(花生、芝麻和棉籽油等)中。其他多不饱和脂肪酸都是由以上4种不饱和脂肪酸衍生而来,通过延长和去饱和作用交替进行来完成的。")
106
①需氧途径 需氧途径由去饱和酶系催化,需O2和NADPH的共同参与。去饱和酶系由去饱和酶及一系列的电子传递体组成。

107
去饱和作用首先发生在饱和脂肪酸的9、10碳原子上,生成单不饱和脂肪酸(如棕榈油酸、油酸)。 动物从该双键向脂肪酸的-COOH端去饱和成多不饱和脂肪酸 植物从该双键向脂肪酸的甲基端去饱和生成如亚油酸、亚麻酸等多烯脂肪酸。
。 动物从该双键向脂肪酸的-COOH端去饱和成多不饱和脂肪酸 植物从该双键向脂肪酸的甲基端去饱和生成如亚油酸、亚麻酸等多烯脂肪酸。")
111
植物的继续去饱和不通过需氧途径,而是在内质网膜上由单不饱和脂肪酸以磷脂或甘油糖脂的形式继续去饱和,也是一个需氧的过程。 由于动物不能合成亚油酸和亚麻酸,但它们对维持其生长十分重要,必须从食物中获得,这些脂肪酸对人类和哺乳动物是必需脂肪酸。但动物能通过去饱和作用和延长脂肪酸碳链的过程将它们转变为二十碳四烯酸。

112
②厌氧途径 厌氧微生物合成单不饱和脂肪酸的方式,它发生在脂肪酸从头合成的过程中。 当FAS系统从头合成到10碳的羟脂酰-ACP(-羟癸酰-ACP)时,由专一性的-羟癸酰-ACP脱水酶催化在、位之间脱水,生成、-烯癸酰-ACP,然后继续参入二碳单位,进行从头合成反应过程,就可产生不同长短的单不饱和脂肪酸。 厌氧途径只能生成单不饱和脂肪酸,因此厌氧微生物中不存在多不饱和脂肪酸。
时,由专一性的-羟癸酰-ACP脱水酶催化在、位之间脱水,生成、-烯癸酰-ACP,然后继续参入二碳单位,进行从头合成反应过程,就可产生不同长短的单不饱和脂肪酸。 厌氧途径只能生成单不饱和脂肪酸,因此厌氧微生物中不存在多不饱和脂肪酸。")
113
三酰甘油的生物合成 由3-磷酸甘油逐步与三分子脂酰CoA缩合生成。 在磷酸甘油脂酰转移酶催化下,3-磷酸甘油与2分子脂酰CoA缩合形成磷脂酸。 在磷酸酶催化下,磷脂酸脱去磷酸生成二酰甘油。 在二酰甘油脂酰转移酶催化下,二酰甘油与1分子脂酰CoA缩合形成三酰甘油。

115
脂肪代谢的调节 脂肪代谢调节与人类健康密切相关,许多疾病如心血管疾病、高血脂和肥胖等都与脂肪代谢的失调有关。 油料作物的出油量也与脂肪代谢的调节有关。脂肪代谢中最重要的部分是脂肪酸的代谢。

116
脂肪酸分解的调节 -氧化是最主要的分解代谢途径,其限速步骤是活化的脂酰CoA从线粒体外转运至线粒体内。 在参与转运的酶及蛋白质中,关键酶是肉毒碱脂酰转移酶I,丙二酸单酰CoA是酶I的抑制剂。 当细胞中能荷较高时,丙二酸单酰CoA含量丰富,使肉毒碱脂酰转移酶I活性降低,脂酰CoA不能穿膜进入线粒体,因而无法氧化放能。

117
当细胞处于高能荷状态时,参与-氧化作用的-羟脂酰CoA脱氢酶被NADH抑制,硫解酶被乙酰CoA抑制。 这些调节方式可以保证细胞在高能荷状态时,抑制脂肪酸的氧化分解放能,而走脂质合成的途径。

118
脂肪酸合成的调节 乙酰CoA羧化酶为变构酶,是脂肪酸合成的限速酶。 在动物体内,乙酰CoA羧化酶由平均约20个单体聚集而成,每个单体兼具生物素羧基载体蛋白、生物素羧化酶和羧基转移酶的功能,但它们只有在聚集状态才具有活性。 柠檬酸能促进无活性的单体聚集成有活性的全酶,从而加速脂肪酸的合成;而软脂酰CoA则相反,它促使聚集物的解体,抑制脂肪酸的合成。

119
当细胞处于高能荷状态时,线粒体中乙酰CoA和ATP含量丰富,可抑制TCA中异柠檬酸脱氢酶的活性,使柠檬酸浓度升高,进入胞质溶胶的柠檬酸可促进乙酰CoA的羧化,同时柠檬酸可裂解成乙酰CoA而参入乙酰CoA的穿梭过程。这些都加速了脂肪酸的合成。

120
当细胞含有过量的脂肪酸时,软脂酰CoA不但抑制乙酰CoA羧化酶的活性,而且还抑制柠檬酸从线粒体基质到胞质溶胶的转移、抑制6-磷酸葡萄糖脱氢酶产生脂肪酸合成所需的还原剂NADPH以及抑制柠檬酸合酶产生柠檬酸从而导致脂肪酸合成的抑制。 因此,当生物体内糖含量高而脂肪酸含量低时,脂肪酸的合成最为有利。

121
3 磷脂的代谢

123
3.1 磷脂的降解 甘油磷脂的降解和甘油三酯相同,先经水解生成甘油、脂肪酸、磷酸及氨基醇,然后水解产物各自按不同的途径进一步分解或转化。 卵磷脂中有4个酯键,需要经过多步水解反应:

124
①磷脂酶催化第一步水解反应 已发现的磷脂酶有磷脂酶A1、磷脂酶A2、磷脂酶C和磷脂酶D等四种,它们对磷脂水解的部位不同,产物也不同。 磷脂酶A1,广泛分布于动物细胞内; 磷脂酶A2存在于蛇毒、蝎毒和蜂毒中; 磷脂酶C存在于动物脑、蛇毒和细菌毒素中; 磷脂酶D主要存在于高等植物中。

126
甘油磷脂的降解 O = CH2O-C-R1 O = - R2-C-O-CH O - = CH2O-P-OCH2CH2N+(CH3)3 -
磷脂酶 甘油磷脂的降解 脂酰甘油磷酸胆碱+脂肪酸 A1、A2、C、D O A1 = CH2O-C-R1 2-脂酰甘油磷酸胆碱+脂肪酸 O A2 = - R2-C-O-CH 1-脂酰甘油磷酸胆碱+脂肪酸 O - = CH2O-P-OCH2CH2N+(CH3)3 - O- C D 二酰甘油+磷酸胆碱 卵磷脂 (磷脂酰胆碱) 磷脂酸 +胆碱 A1:动物体中;A2:蛇、蝎、蜂毒和动物胰脏中 C: 蛇、微生物分泌的毒素、动物脑 D:高等植物中
3. - O- C. D. 二酰甘油+磷酸胆碱. 卵磷脂. (磷脂酰胆碱) 磷脂酸 +胆碱. A1:动物体中;A2:蛇、蝎、蜂毒和动物胰脏中. C: 蛇、微生物分泌的毒素、动物脑. D:高等植物中.")
127
②由磷脂酶A1或磷脂酶A2水解甘油磷脂生成溶血磷脂 溶血磷脂是一类具有较强表面活性的物质,能使红细胞膜和其他细胞膜破坏,引起溶血或细胞坏死。溶血磷脂在溶血磷脂酶作用下,再水解掉一个脂肪酸,生成不具有溶血性的3-甘油磷酸胆碱。

128
③由以上水解酶催化生成的3-甘油磷酸胆碱、磷脂酸、二酰甘油等物质,在磷酸酯酶、脂肪酶等的作用下进一步水解,最终生成脂肪酸、甘油、磷酸及胆碱。 鞘磷脂的降解也需先经历水解过程,再将水解产物分解或转化。

130
磷脂的生物合成 以磷脂酸作前体,需胞嘧啶核苷酸以CDP衍生物形式作活化载体。 有两条途径,以磷酸乙醇胺、磷酸丝氨酸(脑磷脂)为例说明
为例说明")
132
CDP-二酰甘油途径 O CH2O-C-R1 R2-C-O-CH CH2O-P-O-P-O- CH2 O = OH = CH2O-C-R1
CTP PPi - - = R2-C-O-CH O O 胞苷酰转移酶 胞嘧啶 - - = = = CH2O-P-OH - - - OH OH O 磷脂酸 OH CDP-二酰甘油 Ser CMP O PE CH2O-C-R1 R2-C-O-CH CH2O-P-O-CH2-C- COOH O = OH 磷脂酰丝氨酸脱羧酶 = CH2OCR1 O CO2 PS NH2 - - = R2COCH O - - = - = CH2O-P-O- CH2CH2NH2 - - - OH H

133
利用CDP-乙醇胺的途径 HOCH2CH2NH2 乙醇胺 磷脂酸 ATP H2O O ADP = Pi HO-P-O-CH2CH2NH2
磷酸酶 O ADP = Pi HO-P-O-CH2CH2NH2 磷酸乙醇胺 O = - CH2O-C-R1 OH CTP O - = PPi R2-C-O-CH - 胞嘧啶 O O CH2OH = = CH2O-P-O-P-O- CH2CH2NH2 O 二酰甘油 OH - - OH OH 磷脂酰乙醇胺 PE CDP-乙醇胺 CMP

136
体内磷脂酰丝氨酸合成是通过Ca2+激活的酰基交换反应生成,由磷脂酰乙醇胺与丝氨酸反应生成磷脂酰丝氨酸和乙醇胺。

137
鞘磷脂的生物合成 鞘磷脂与甘油磷脂在结构上的不同之处在于由鞘氨醇代替了甘油,它是与1分子脂肪酸、磷酸和胆碱结合而形成的。

138
鞘磷脂的合成分为3个阶段: ①鞘氨醇的合成:由软脂酰CoA与丝氨酸经一系列酶促反应形成。

139
②神经酰胺的形成:由鞘氨醇的氨基与脂酰CoA的脂酰基连结形成。

140
③鞘磷脂的合成由神经酰胺接受CDP-胆碱上的磷酸胆碱形成。

141
3.2 糖脂的降解与生物合成 3.2.1 糖脂的降解 糖苷酶可将糖脂上的糖基水解下来,其他的成分在各种脂酶的作用下可水解成甘油或鞘氨醇、脂肪酸等。 例如,当植物叶细胞受到破坏时,单半乳糖二脂酰甘油(MGDG)和双半乳糖二脂酰甘油(DGDG)可在半乳糖脂酶、-半乳糖苷酶等酶的催化下,迅速水解成甘油、脂肪酸和半乳糖。
和双半乳糖二脂酰甘油(DGDG)可在半乳糖脂酶、-半乳糖苷酶等酶的催化下,迅速水解成甘油、脂肪酸和半乳糖。")
142
糖脂的生物合成 ⑴甘油糖脂的生物合成 植物体内甘油糖脂主要有单半乳糖二脂酰甘油和双半乳糖二脂酰甘油,它们是叶绿体膜中的主要脂类,它们的合成是在叶绿体被膜上进行的。

143
①单半乳糖二脂酰甘油(MGDG)的合成 先合成的磷脂酸水解掉磷酸生成二脂酰甘油。在UDP-半乳糖-二脂酰甘油半乳糖基转移酶催化下,二脂酰甘油接受UDP-半乳糖上的半乳糖基生成MGDG。
的合成 先合成的磷脂酸水解掉磷酸生成二脂酰甘油。在UDP-半乳糖-二脂酰甘油半乳糖基转移酶催化下,二脂酰甘油接受UDP-半乳糖上的半乳糖基生成MGDG。")
144
②双半乳糖二脂酰甘油(DGDG)的合成 单半乳糖二脂酰甘油再接受一分子UDP-半乳糖上的半乳糖基,即可生成DGDG。 植物体内合成多烯脂肪酸时,去饱和酶的底物不是来自脂肪酸,而是磷脂或甘油糖脂,如MGDG,其脂酰基R2可以被去饱和酶作用继续脱饱和形成多烯脂肪酸。
的合成 单半乳糖二脂酰甘油再接受一分子UDP-半乳糖上的半乳糖基,即可生成DGDG。 植物体内合成多烯脂肪酸时,去饱和酶的底物不是来自脂肪酸,而是磷脂或甘油糖脂,如MGDG,其脂酰基R2可以被去饱和酶作用继续脱饱和形成多烯脂肪酸。")
145
⑵鞘糖脂的生物合成 鞘糖脂生物合成的前体物质也是神经酰胺。神经酰胺的末端羟基接受UDP-糖上的糖基,即可生成脑苷脂。
如果在脑苷脂的糖基上继续添加糖基或其它基团,可形成其它鞘糖脂,如神经节糖脂。 脑苷脂的合成还可以鞘氨醇为前体,先接受糖基形成鞘氨醇糖苷然后再脂酰化而完成其合成。

146
4 胆固醇的代谢

147
4.1 胆固醇的生物合成 场所:肝脏,占全身合成总量的3/4。(小肠10%,皮肤、肾上腺皮质、性腺等也能合成)
4.1 胆固醇的生物合成 场所:肝脏,占全身合成总量的3/4。(小肠10%,皮肤、肾上腺皮质、性腺等也能合成) 合成的胆固醇以脂蛋白的形式快速输送到血液中。 转变和消耗:每日合成约1-1.5克,其中约0.8克转变为胆酸和脱氧胆酸。胆汁中的胆酸盐经胆管输入十二指肠,参与脂类消化。每日随粪便以粪固醇排出约0.4克。 碳源和原料:乙酰CoA、NADPH、ATP。
 合成的胆固醇以脂蛋白的形式快速输送到血液中。 转变和消耗:每日合成约1-1.5克,其中约0.8克转变为胆酸和脱氧胆酸。胆汁中的胆酸盐经胆管输入十二指肠,参与脂类消化。每日随粪便以粪固醇排出约0.4克。 碳源和原料:乙酰CoA、NADPH、ATP。")
148
①由3分子乙酰CoA合成1分子3-甲基-3,5-二羟戊酸(甲瓦龙酸,MVA) 2分子乙酰CoA缩合成乙酰乙酰CoA后再与1分子乙酰CoA缩合成-羟--甲基戊二酸单酰CoA(HMG-CoA)。
 2分子乙酰CoA缩合成乙酰乙酰CoA后再与1分子乙酰CoA缩合成-羟--甲基戊二酸单酰CoA(HMG-CoA)。")
149
HMG-CoA是酮体合成与胆固醇合成的分支点。若被裂解酶作用,则合成酮体乙酰乙酸;若被还原酶作用,则被NADPH还原成MVA。 此反应也是胆固醇合成的限速反应。

150
②由MVA生成异戊烯醇焦磷酸酯(IPP) MVA经3次磷酸化生成的3-磷酸-5-焦磷酸MVA,又在脱羧酶作用下脱羧而形成IPP。 IPP不仅是合成胆固醇的前体,也是植物合成萜类物质、昆虫合成保幼激素、蜕皮素等的活泼前体。
 MVA经3次磷酸化生成的3-磷酸-5-焦磷酸MVA,又在脱羧酶作用下脱羧而形成IPP。 IPP不仅是合成胆固醇的前体,也是植物合成萜类物质、昆虫合成保幼激素、蜕皮素等的活泼前体。")
151
(3)由6分子IPP缩合成1分子鲨烯(squalene) 1分子IPP先异构成3,3-二甲基丙烯焦磷酸酯(DPP),然后与2分子IPP逐一进行头尾缩合,先后生成牻牛儿焦磷酸酯(GPP)和法呢焦磷酸酯(FPP)。 牻牛儿焦磷酸酯可进一步缩合成二萜及多萜类,在植物体内可进一步转化为叶黄素、番茄红素、胡萝卜素等。
由6分子IPP缩合成1分子鲨烯(squalene) 1分子IPP先异构成3,3-二甲基丙烯焦磷酸酯(DPP),然后与2分子IPP逐一进行头尾缩合,先后生成牻牛儿焦磷酸酯(GPP)和法呢焦磷酸酯(FPP)。 牻牛儿焦磷酸酯可进一步缩合成二萜及多萜类,在植物体内可进一步转化为叶黄素、番茄红素、胡萝卜素等。")
153
2分子FPP尾尾缩合并被NADPH还原脱去2分子焦磷酸生成鲨烯。

154
④由鲨烯生成胆固醇 鲨烯在2、3位上环氧化生成2,3-环氧鲨烯;然后整条长链环化形成羊毛固醇。 羊毛固醇经3次脱甲基,双键从7、8位至5、6位的移动以及侧链双键被NADPH还原成单键等多步反应,最终形成胆固醇。

155
环氧鲨烯除了在动物中形成胆固醇外,在植物中可转化成豆甾醇,在真菌中可转化成麦角固醇。 胆固醇主要在脊椎动物的肝脏中合成。在胆固醇合成的4个阶段中,第1阶段由膜酶催化;第2、3阶段由可溶性酶催化;第4阶段即合成鲨烯后,鲨烯被固体载体蛋白(SCP)转运至内质网膜上继续进行合成反应。
转运至内质网膜上继续进行合成反应。")
156
胆固醇的转化 胆固醇在动物体内不仅可以在C3的羟基上接受脂酰CoA的脂酰基而酯化成胆固醇脂,还可转化成具有重要生理功能的物质,如胆酸、类固醇激素、维生素D等。

157
①转化为胆酸及其衍生物 胆固醇在羟化酶及脱氢酶的催化下,在C7、C12位上发生羟基化,侧链C24位氧化成羧酸,从而转变为胆酸。 胆酸在消耗ATP的条件下可形成胆酰CoA,胆酰CoA与牛磺酸(H2NCH2CH2SO3H)或甘氨酸缩合形成牛磺胆酸或甘氨胆酸,这两种胆汁盐对油脂的消化和脂溶性维生素的吸收有重要作用。
或甘氨酸缩合形成牛磺胆酸或甘氨胆酸,这两种胆汁盐对油脂的消化和脂溶性维生素的吸收有重要作用。")
158
②转化为类固醇激素 胆固醇在羟化酶、脱氢酶、异构酶和裂解酶的催化下,可转化成各种类固醇激素,如糖皮质激素、盐皮质激素、孕酮、雄激素和雌激素等。

159
③转化为维生素D 胆固醇先转化成7-脱氢胆固醇,后者在紫外光作用下,C9与C10间发生开环,再进一步转化变为维生素D3。 麦角固醇在紫外线作用下,也可转变成维生素D2。

162
5 脂类代谢紊乱

163
4.1 酮血症、酮尿症 当人体患有糖尿病,糖类物质利用受阻或长期不能进食,机体所需能量不能从糖的氧化取得,于是大量动用脂肪提供能量,脂肪酸大量氧化,生成的酮体超过了肝外组织所能利用的限度,导致血液中酮体堆积,含量升高,临床上称为酮血症。

164
5.2 脂肪肝 肝脏在脂类代谢中的作用特别重要,能合成脂蛋白,利于脂类运输;是脂肪酸氧化和酮体形成的主要场所
5.2 脂肪肝 肝脏在脂类代谢中的作用特别重要,能合成脂蛋白,利于脂类运输;是脂肪酸氧化和酮体形成的主要场所 肝的脂类含量并不多(约为4%),主要是磷脂,脂肪仅占脂类总量的1/4。 脂肪肝是当肝脏脂蛋白不能及时将肝细胞脂肪运出,造成脂肪在肝细胞中的堆积所致。脂肪肝患者的肝脏脂肪含量竟超过10%,
,主要是磷脂,脂肪仅占脂类总量的1/4。 脂肪肝是当肝脏脂蛋白不能及时将肝细胞脂肪运出,造成脂肪在肝细胞中的堆积所致。脂肪肝患者的肝脏脂肪含量竟超过10%,")


























 不饱和脂肪酸的氧化 酮体 磷脂的代谢 脂肪酸代谢的调控.>")
 信号传导:激素、胞内信使.>")

 反应部位 器官,细胞内定位 生理意义 代谢调节 主要调节点,主要变构抑制剂、变构激活剂 各代谢途径之间的联系和调控.>")
:核蛋白体组成成分 转移 RNA ( tRNA ):转运氨基酸 信使 RNA ( mRNA ):蛋白质合成模板 不均一核 RNA ( hnRNA ):成熟 mRNA 的前体 小核 RNA ( snRNA ):>")



.>")


物质在生物体的分解及合成代谢。要求学生重点掌握脂肪酸在生物体内的氧化分解途径—β-氧化和从头合成途径,了解脂类物质的功能和其他的氧化分解途径。 思考 脂类代谢 返回.>")







