Download presentation
0
糖及糖代谢 Carbohydrates & Carbohydrates Metabolism
宋潇达

1
目录 Section 1. 总览 Section 2. 糖类和糖生物学 Section 3. 糖代谢 Section 4. 糖原代谢
糖酵解 三羧酸循环 戊糖磷酸途径 糖异生 好复杂,我看也醉了。不用担心! 目录

2
第一部分. 总览 细胞膜 三羧酸循环 NADH ATP 1953诺贝尔奖 生物氧化 线粒体膜 淀粉 乙酰辅酶A 核糖-5-磷酸 葡萄糖
蛋白酶消化 乙酰辅酶A 核糖-5-磷酸 葡萄糖 吸收 戊糖磷酸途径 丙酮酸 乙醇 1907诺贝尔奖 1929诺贝尔奖 糖异生 糖酵解 非糖前体 乳酸 丙酮酸 甘油 葡萄糖 乳酸 简要介绍糖类代谢这章的主要内容,理清思路。以葡萄糖为中心,三个去路,三个来源。 分解 合成 1970诺贝尔奖 1902诺贝尔奖 1937诺贝尔奖 糖原

3
第二部分. 糖类与糖生物学 糖类: 生物能量的载体 氨基酸和核酸的生物合成前体 蛋白在细胞内的锚定和信号传递部分 结构支撑:细胞壁和外骨骼
快速讲解糖类的四种生物学功能,板书“能量,合成前体,信号,结构” 例子:决定了人的血型

4
第二部分. 糖类与糖生物学 糖类: 多羟基醛,酮及其聚合物和衍生物总称 一般结构式: (CH2O)n 单糖 1 unit
寡糖 units 多聚糖 >20 units 三种主要类别 1-19 >19 二糖: 2 个糖单位 后缀 “-ose” 糖连接物(Glycoconjugates): 糖通过共价键连接到蛋白或者脂类形成的化合物 讲解糖类的定义:糖定义为多羟基醛、酮及其缩聚物和某些衍生物。有单糖、寡糖、多糖和复合糖类。强调单糖,二糖。单糖是糖结构的单体,可用一个经验公式(CH2O)n 表示,但有意外。让同学回去自己看书上单糖,那些不是,比如鼠李糖, 脱氧核糖 C5H10O4之类的。 n
: 糖通过共价键连接到蛋白或者脂类形成的化合物. 讲解糖类的定义:糖定义为多羟基醛、酮及其缩聚物和某些衍生物。有单糖、寡糖、多糖和复合糖类。强调单糖,二糖。单糖是糖结构的单体,可用一个经验公式(CH2O)n 表示,但有意外。让同学回去自己看书上单糖,那些不是,比如鼠李糖, 脱氧核糖 C5H10O4之类的。 n.")
5
Tips:生物体内一般是D型糖,L型氨基酸
典型单糖的Fischer 投影式 Tips:生物体内一般是D型糖,L型氨基酸 3-碳醛糖 3-碳酮糖 Chiral center L(S) Glyceraldehyde 甘油醛 D(R) Glyceraldehyde 甘油醛 Dihydroxyacetone (no chiral center) 二羟丙酮 4-碳醛糖 New Chiral center 介绍糖类的分类,一般分为醛糖和酮糖两类。最简单的三碳糖是甘油醛和二羟基丙酮。Fischer投影式醛糖中氧化数最高的碳原子指定为C-1,酮糖中氧化数最高的碳原子指定为C-2,除最简单的二羟丙酮外,都是手性分子。醛糖中手性碳的数目为n-2,异构体的数目为2n-2。糖的构型有D型和L型。D型糖是指具有最高编号的手性碳,即离羰基碳最远的手性碳连接的- OH在Fischer投影式中是朝向右的。介绍生物体内一般都是D型糖,L型氨基酸。介绍糖可以从起初两个三碳糖衍生化下来,介绍赤藓糖,苏阿糖回忆有机的命名赤和苏。 D/L-构型 距羰基最远的手性碳上羟基向右的 为D型,向左为L型 α/β构型 C1 上羟基在碳链右侧的称 α 型, 在左侧的称为β型 Erythrose 赤藓糖 Threose 苏阿糖 Same chiral center as D(R) Glyceraldehyde
 Glyceraldehyde. 甘油醛. D(R) Glyceraldehyde. 甘油醛. Dihydroxyacetone. (no chiral center) 二羟丙酮. 4-碳醛糖. New Chiral center. 介绍糖类的分类,一般分为醛糖和酮糖两类。最简单的三碳糖是甘油醛和二羟基丙酮。Fischer投影式醛糖中氧化数最高的碳原子指定为C-1,酮糖中氧化数最高的碳原子指定为C-2,除最简单的二羟丙酮外,都是手性分子。醛糖中手性碳的数目为n-2,异构体的数目为2n-2。糖的构型有D型和L型。D型糖是指具有最高编号的手性碳,即离羰基碳最远的手性碳连接的- OH在Fischer投影式中是朝向右的。介绍生物体内一般都是D型糖,L型氨基酸。介绍糖可以从起初两个三碳糖衍生化下来,介绍赤藓糖,苏阿糖回忆有机的命名赤和苏。 D/L-构型 距羰基最远的手性碳上羟基向右的. 为D型,向左为L型. α/β构型 C1 上羟基在碳链右侧的称 α 型, 在左侧的称为β型. Erythrose. 赤藓糖. Threose. 苏阿糖. Same chiral center. as D(R) Glyceraldehyde.")
6
差向异构体 epimer:只有一个碳原子构型不同的两种糖
Erythrose Threose 5-碳醛糖 Ribose 核糖 Arabinose Xylose Lylose 6-碳醛糖 6-碳酮糖 跳到第6页,衍生到五碳糖。要同学记住核糖,脱氧核糖,记忆两者区别:以脱衬衫为例,脱氧核糖缺第一个氧(扣子)。衍生到六碳糖,记住葡萄糖,果糖。引出概念差向异构体。 Allose Altrose Glucose 葡糖糖 Mannose 甘露糖 Gulose Idose Galactose Talose Fructose 果糖 差向异构体 epimer:只有一个碳原子构型不同的两种糖
。衍生到六碳糖,记住葡萄糖,果糖。引出概念差向异构体。 Allose. Altrose. Glucose. 葡糖糖. Mannose. 甘露糖. Gulose. Idose. Galactose. Talose. Fructose. 果糖. 差向异构体 epimer:只有一个碳原子构型不同的两种糖.")
7
1902年对嘌呤和糖类的合成研究被授予1902年诺贝尔化学奖 Hermann Emil Fischer 赫尔曼·埃米尔·费歇尔
1852年出生于德国 1872年合成了苯肼 1883年首先研究了葡萄糖的性质 葡萄糖被氧化为葡萄糖酸 葡萄糖被还原为醇 葡萄糖与苯肼的反应生成苯腙 1891年提出Fischer投影式 Glucose 1902年对嘌呤和糖类的合成研究被授予1902年诺贝尔化学奖 Hermann Emil Fischer 赫尔曼·埃米尔·费歇尔 葡萄糖的检测(例如糖尿病人血液和尿液中葡萄糖含量的检测?) 过去:菲林试剂,测定消耗了多少氧化剂 现在:葡萄糖氧化酶,用过氧化酶催化H2O2和无色底物反应生成有色物质 直接说Fisher对葡萄糖做了多种研究,指着葡萄糖结构快速讲化学反应,醛基—氧化还原,成腙成糖脎,高碘酸氧化。举例:葡萄糖的检测,糖尿病在以往都是采用菲林试剂,现在用酶法检测。 葡萄糖氧化酶 葡萄糖 O2 葡萄酸-δ-内酯 H2O2
 过去:菲林试剂,测定消耗了多少氧化剂. 现在:葡萄糖氧化酶,用过氧化酶催化H2O2和无色底物反应生成有色物质. 直接说Fisher对葡萄糖做了多种研究,指着葡萄糖结构快速讲化学反应,醛基—氧化还原,成腙成糖脎,高碘酸氧化。举例:葡萄糖的检测,糖尿病在以往都是采用菲林试剂,现在用酶法检测。 葡萄糖氧化酶. 葡萄糖. O2. 葡萄酸-δ-内酯. H2O2.")
8
首次用硫酸二甲酯对糖类进行O-甲基化来研究糖类结构 提出了Haworth透视式 在碳水化合物和维生素上的研究于1937年获得诺贝尔化学奖
1883年出生于英国 首次用硫酸二甲酯对糖类进行O-甲基化来研究糖类结构 提出了Haworth透视式 在碳水化合物和维生素上的研究于1937年获得诺贝尔化学奖 Sir Walter N Haworth β-D-吡喃葡萄糖 变旋 醛糖和酮糖可以形成环式的半缩醛。有5员环或6员环结构,称为呋喃糖或吡喃糖。一般是以吡喃糖为主,让学生看反应有两个结构。关注1号位的碳,这个碳是手性,称异头碳。有α、β两个新异构体。它们不同的环式和开链式处于平衡中。椅式构象比船式更稳定。D/L-构型 环上C按顺时针排列,环上编号最大手性C的羟甲基(若无羟甲基,则以该C 上的H) 在面上为 D,面下为 L型α/β构型 半缩醛 (酮 ) 羟基与环上编号最大的手性C的羟甲基(若无羟甲基,则以该C 上的H) 处于同侧为 β 型,如果处于异侧,则为α 型。 α-D-吡喃葡萄糖
 在面上为 D,面下为 L型α/β构型 半缩醛 (酮 ) 羟基与环上编号最大的手性C的羟甲基(若无羟甲基,则以该C 上的H) 处于同侧为 β 型,如果处于异侧,则为α 型。 α-D-吡喃葡萄糖.")
9
二糖介绍 结构 单糖种类 糖苷键类型 还原性 麦芽糖 α-D-葡萄糖 α-1,4-糖苷键 有 乳糖 α-D-半乳糖 β-D-葡萄糖
β-1,4-糖苷键 蔗糖 β-D-果糖 α-1,2-糖苷键 无 介绍单糖可以通过糖苷键形成寡糖和多糖 3种重要的双糖有麦芽糖(α-1,4)、乳糖和蔗糖。如何看还原性,加深环式和开链式处于平衡的状态 未水解的乳糖人体不能利用,势必由肾脏排出而出现在尿中。如果缺少乳糖酶,饮乳后会引起腹胀、恶心、呕吐、水泻等症状。
、乳糖和蔗糖。如何看还原性,加深环式和开链式处于平衡的状态. 未水解的乳糖人体不能利用,势必由肾脏排出而出现在尿中。如果缺少乳糖酶,饮乳后会引起腹胀、恶心、呕吐、水泻等症状。")
10
请自己总结:连接键,分子形状,分子量大小,遇碘显色情况
多糖及糖缀合物介绍 多糖,淀粉,糖原,葡聚糖,纤维素 请自己总结:连接键,分子形状,分子量大小,遇碘显色情况 琼脂:果冻状物质,实验上用途 agar:用作培养基平板的 agarose:用DNA分离纯化 透明质酸:玻尿酸, 肝素:抗凝血药 硫酸软骨素:降血脂抗凝血 细菌酯多糖:内毒素,会引起人体过敏发热,严重导致死亡 多糖部分让学生自己回去总结,举例在实验中误用agar和agarose的情况,以及多加了agarose培养基长不出来

11
淀粉消化主要在小肠内进行。糖被消化成单糖后才能在小肠被吸收,再经门静脉进入肝脏。
糖类化合物的消化和吸收: 多糖类 淀粉 糊精 寡糖 麦芽糖 葡萄糖 淀粉酶 唾液淀粉酶 糊精酶 主动吸收 淀粉 口腔 胃 小肠 葡萄糖 血液 麦芽糖酶 Na+依赖型葡萄糖转运体 淀粉消化主要在小肠内进行。糖被消化成单糖后才能在小肠被吸收,再经门静脉进入肝脏。 双糖及寡糖类 糖的降解注意是全部降解为单糖吸收。人的消化道无水解纤维素的酶。 提问葡萄糖是如何具体经过吸收进入细胞,血液? 蔗糖酶 葡萄糖是如何具体吸收通过细胞进入血液? ----- 见后续糖酵解部分 蔗糖 葡萄糖,果糖 乳糖酶 乳糖 葡萄糖,半乳糖 海藻糖酶 海藻糖 葡萄糖

12
食物中的糖类 消化 葡萄糖 吸收 血糖 运输 组织细胞 (葡萄糖) 氧化分解 合成 分解 转变 CO2+H2O+能量 肝糖元 肌糖元 脂肪、某些氨基酸等
 氧化分解 合成 分解 转变 CO2+H2O+能量 肝糖元 肌糖元 脂肪、某些氨基酸等")
13
第三部分. 糖代谢 3.1 糖酵解 3.2 三羧酸循环 3.3 戊糖磷酸途径 3.4 糖异生 糖酵解应掌握: 总反应及计算 限速反应
三种抑制剂及机理 底物水平磷酸化 主要的调控 介绍糖代谢,需要了解总反应,限速反应,抑制剂,磷酸化,调控机制。 有机和实验是基础,分子结构不用记忆,但很重要,帮助理解。 需要多思考为什么反应会发生,有什么意义?而不是直接结果。这是药大教学和其他高水平大学差距。以自己为例虽然拿个好看分数但到头还得重新自学。提醒学生要打起十二分精神。 化学结构很重要,有助于理解反应机理,但在这里不要记忆。 建议记忆几个关键的结构。

14
The Embden-Meyerhof (Warburg) Pathway
3.1 糖酵解 细胞溶质 Glucose ATP The Embden-Meyerhof (Warburg) Pathway 所有细胞都产生糖酵解 细胞内位置: 细胞溶质 不同的细胞- 糖酵解速率不一样 总共十个反应 葡萄糖 (输入物质) 两个丙酮酸(输出物质), 没有碳原子的减少 消耗一个葡萄糖分子产生两个ATP , 两个 NADH ATP Phase I 投入部分 -2ATP NADH G-3-P NADH ATP Phase II 收获部分 +4ATP, 2NADH ATP 总述糖酵解,所有生物细胞都存在,细胞内位置: 细胞溶质。不需要氧,包括没有线粒体的血红细胞。不同的细胞- 糖酵解速率不一样。输入物质,葡萄糖,输出两个丙酮酸(输出物质), 没有碳原子的减少。 10个反应,分为两步,到甘油三磷酸时为投入部分,-2ATP,后面到丙酮酸为收获部分,+4ATP,2NADH ATP ATP 丙酮酸
 Pathway. 所有细胞都产生糖酵解. 细胞内位置: 细胞溶质. 不同的细胞- 糖酵解速率不一样. 总共十个反应. 葡萄糖 (输入物质) 两个丙酮酸(输出物质), 没有碳原子的减少. 消耗一个葡萄糖分子产生两个ATP , 两个 NADH. ATP. Phase I. 投入部分. -2ATP. NADH. G-3-P. NADH. ATP. Phase II. 收获部分. +4ATP, 2NADH. ATP. 总述糖酵解,所有生物细胞都存在,细胞内位置: 细胞溶质。不需要氧,包括没有线粒体的血红细胞。不同的细胞- 糖酵解速率不一样。输入物质,葡萄糖,输出两个丙酮酸(输出物质), 没有碳原子的减少。 10个反应,分为两步,到甘油三磷酸时为投入部分,-2ATP,后面到丙酮酸为收获部分,+4ATP,2NADH. ATP. ATP. 丙酮酸.")
15
糖酵解的反应 ATP ADP ATP H2O ADP ADP ATP
转到下一页糖酵解的分子结构图,简要描述下过程。学生猜下哪步很重要,调控步?一般而言单箭头,不可逆。下面我将详细介绍每个反应(也许不会考但很重要),最后总结。 ATP ADP NADH, H+ NAD+, Pi
,最后总结。 ATP. ADP. NADH, H+ NAD+, Pi.")
16
Glycolysis-Reaction 1: phosphorylation of glucose
ATP ADP Mg2+ 己糖激酶 葡萄糖 6-磷酸葡萄糖 ΔG’°= kJ/mol ⌘ 己糖激酶 or 葡萄糖激酶 ⌘ Priming reaction – ATP 在这里被消耗,是为了更好的获得 ⌘ ATP的消耗使葡萄糖的磷酸化不可逆 第一步反应,葡萄糖在己糖激酶或葡萄糖激酶的作用下在6位引入磷酸基团。从吉布斯自由能看反应向右进行。

17
Glycolysis-Reaction 1: phosphorylation of glucose
ATP ADP Mg2+ 己糖激酶 葡萄糖 6-磷酸葡萄糖 ΔG’°= kJ/mol 1. 镁离子有什么作用? 2. 为什么ATP不先和水反应而和羟基反应? 3. 这个反应的背后有什么意义? 同学们思考下:1.镁离子有什么作用?2.羟基和水(板书HOH)水的酸性强,为什么ATP不先和水反应而和羟基,这不科学! 3.为什么要发生这个反应,有什么意义。我看有些同学在翻书,大家不用翻书,书上没有,大家带着问题听我讲。Mg离子在生物体内很重要,板书如下结构 Mg离子封闭了磷酸根离子的氧负离子使磷原子更容易被亲核进攻。氟化物容易与镁形成络合物,有毒性。
水的酸性强,为什么ATP不先和水反应而和羟基,这不科学! 3.为什么要发生这个反应,有什么意义。我看有些同学在翻书,大家不用翻书,书上没有,大家带着问题听我讲。Mg离子在生物体内很重要,板书如下结构 Mg离子封闭了磷酸根离子的氧负离子使磷原子更容易被亲核进攻。氟化物容易与镁形成络合物,有毒性。")
18
诱导契合:己糖激酶绑定葡萄糖后,导致大规模的构象变化
己糖激酶的诱导契合模型 己糖激酶绑定葡萄糖后,导致大规模的构象变化,将水分子挤出去,产生一个疏水的环境。ATP只能与葡萄糖反应。 PDB: 2YHX PDB: 1HKJ 诱导契合:己糖激酶绑定葡萄糖后,导致大规模的构象变化

19
己糖激酶的诱导契合:木糖的欺骗 葡萄糖 木糖
ATP ATP 葡萄糖 木糖 木糖代替葡萄糖,结果发现己糖激酶能够催化ATP水解。很可能是因为体积较小的木糖进入活性中心以后,无法赶走“水分子”,诱使酶“上当受骗”而将ATP的γ-磷酸转移给留在活性中心的水分子上的羟基所致。 木糖代替葡萄糖,结果发现己糖激酶能够催化ATP水解,诱使酶“上当受骗”。 因此木糖类常作为糖尿病人的甜味剂。

20
糖 甜度 果糖 蔗糖 葡萄糖 木糖 糖精 甜菊苷 173.3 100 74.3 40 50000 20000 鼠李糖 麦芽糖 半乳糖 棉子糖
阿斯巴甜 32.5 32.1 22. 6 16.1 15000 糖精 阿斯巴甜 甜菊苷

21
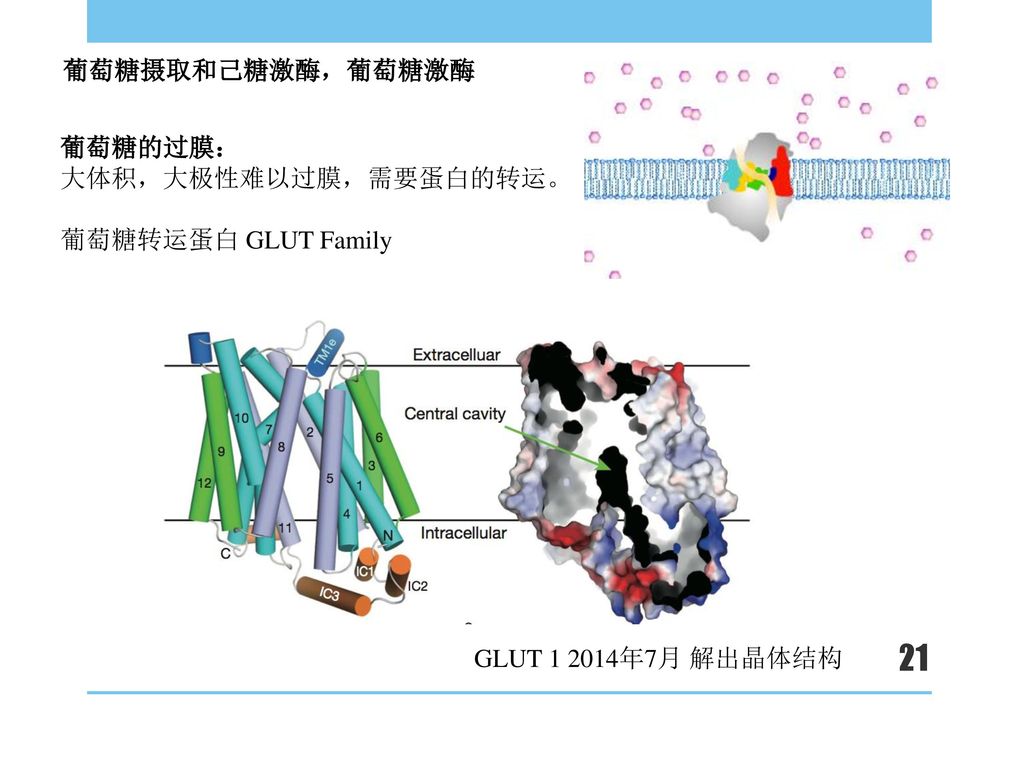
大体积,大极性难以过膜,需要蛋白的转运。 葡萄糖转运蛋白 GLUT Family
葡萄糖摄取和己糖激酶,葡萄糖激酶 葡萄糖的过膜: 大体积,大极性难以过膜,需要蛋白的转运。 葡萄糖转运蛋白 GLUT Family 葡萄糖为大极性大体积的分子,难以透过膜,在膜间转运需要转运蛋白,GLUT 1在今年被解出晶体结构,大家可以看到有个cavity,通道。板书画葡萄糖的自由能=RTln(C细胞外/细胞内) GLUT 年7月 解出晶体结构
 GLUT 年7月 解出晶体结构.")
23
己糖激酶和葡萄糖激酶的比较 葡萄糖摄取和己糖激酶,葡萄糖激酶 6-磷酸葡萄糖体积,极性大于葡萄糖,因此不能扩散到细胞外。 己糖激酶
存在 几乎所有的细胞 肝细胞 底物特异性 葡萄糖、甘露糖、氨基葡萄糖、 果糖、2-脱氧葡萄糖等己糖 葡萄糖和2-脱氧葡萄糖 对葡萄糖的Km 0.1mM 10mM 产物反馈抑制 G-6-P反馈抑制 不受G-6-P反馈抑制 基因表达 组成酶 诱导酶 葡萄糖在体内被磷酸化,极性增大,体积也增加。就不能通过转运蛋白的孔。会被截留在细胞内。随着细胞内葡萄糖的转化,浓度差促使胞外葡萄糖进入细胞。 己糖激酶和葡萄糖激酶的比较,意义在于在餐后,血液内大量葡糖糖时,快速摄取入肝。在血液中4mM葡萄糖时,葡萄糖激酶被激活。

24
Glycolysis-Reaction 2: Conversion of Glucose 6-phosphate
to Fructose 6-Phosphate Mg2+ 磷酸己糖异构酶 6-磷酸葡萄糖 6-磷酸果糖 ΔG’°= 1.7 kJ/mol ⌘ 反应的意义 下一步反应的1号位磷酸化,伯醇的活性高于仲醇 ⌘ 烯-二醇中间体 (Ene-diol )
")
25
磷酸己糖异构酶 Binding and Opening of the ring
Dissociating and Closing of the ring 磷酸己糖异构酶 (Glu)BH (Glu)B (Glu)B H+ H+ Cis-enediol formation Acid catalysis
BH. (Glu)B. (Glu)B. H+ H+ Cis-enediol formation. Acid catalysis.")
26
Reaction 3: Phosphorylation of Fructose 6-phosphate to Fructose 1,6-Bisphosphat
ATP ADP Mg2+ 磷酸果糖激酶-1 6-磷酸果糖 1,6-二磷酸果糖 ΔG’°= kJ/mol ⌘ 加入第二个磷酸基团为关键的调控步骤 ⌘ 关键步骤,较高的ΔG 意味着磷酸果糖激酶被高度调控

27
磷酸果糖激酶 糖结合位点 ATP,Mg离子 结合位点 ATP,ADP竞争结合位点 ADP未结合,无活性 ADP结合后,有活性

28
Reaction 4: Aldolase ⌘ 分为两类,动物为Class I 醛缩酶
1,6-二磷酸果糖 3-磷酸甘油醛 磷酸二羟丙酮 ΔG’°= 23.8 kJ/mol ⌘ 分为两类,动物为Class I 醛缩酶 ⌘ Class I 醛缩酶在活性位点Lys形成西弗碱Schiff base中间体 ⌘ 如何证明Schiff base中间体?

29
Class I aldolases H2O Class II aldolases

30
Class I aldolases 229 Lys 含有四个亚基 每个亚基都有催化活性 229Lys 形成Schiff碱 PDB: 1J4E

31
Class II aldolases Zn2+ Zn2+ 二聚体 催化中心有两个锌离子 PDB: 1ZEN

32
Reaction 5: Interconversion of the Triose Phosphates
磷酸丙糖异构酶 磷酸二羟丙酮 3-磷酸甘油醛 ΔG’°= 7.5 kJ/mol ⌘ 烯-二醇中间体 (Ene-diol ) ⌘ 磷酸丙糖异构酶具有特殊αβ barrel结构
 ⌘ 磷酸丙糖异构酶具有特殊αβ barrel结构.")
33
与一般酶不同,起始Glu作碱

34
底物 95 His 165 Glu PDB: 2YPI

35
TIM: αβ barrel GFP: β barrel ★Triosephosphate Isomerase 结构是 αβ barrel, 也称为 TIM barrel。 ★αβ barrel 有8 α-helices围绕着8 parallel β-strands 。 ★β-strands 和 α-helices之间有loop相连。 ★TIM barrel 结构中 loop 形成了一个盖子,盖在活性位点上部。

36
与过渡态相似的化合物(Analogs) 过渡态 B. 2-phosphoglycolate A. 2-phosphoglycohydroxamate 过渡态类似物 Phosphoglycohydroxamate (A) 和 2-phosphoglycolate (B) 结合 TIM 155 and 100 times
 和 2-phosphoglycolate (B) 结合 TIM 155 and 100 times.")
37
Glycolysis Phase II 收获部分
ADP ATP NAD+, Pi NADH, H+ H2O ADP ATP ⌘ 代谢过程中共产生4ATP,2NADH ⌘ 糖酵解净ATP产量为2ATP ⌘ 有两个高能磷酸中间体 1,3-二磷酸甘油 磷酸烯醇丙酮酸

38
Reaction 6: Oxidation of Glyceraldehyde 3-Phosphate to 1,3-Bisphosphoglycerate
NAD+ NADH+H+ 3-磷酸甘油醛脱氢酶 3-磷酸甘油醛 1,3-二磷酸甘油酸 磷酸 ΔG’°= 6.3 kJ/mol ⌘ Gly-3P 被氧化为1,3-BPG ⌘ 醛与酶中半胱氨酸巯基成硫酯中间体,共价键四面体过渡态 ⌘ 抑制剂:碘乙酸,砷酸

39
反应机理:

40
碘乙酸,砷酸的抑制原理 I. 碘乙酸与催化中心巯基共价结合使酶失活 II. 砷酸的结构与磷酸类似,生成类似1-arseno-3-phosphoglycerate 砷酸酯容易水解,导致后续无法产生ATP

41
Reaction 7: Phosphoryl Transfer from 1,3-Bisphosphoglycerate to ADP
Mg2+ 磷酸甘油酸激酶 1,3-二磷酸甘油酸 3-磷酸甘油酸 ΔG’°= kJ/mol ⌘ ATP 从高能磷酸底物中生成 ⌘ 底物磷酸化 "substrate-level phosphorylation” ⌘ 红细胞内会出现生成2,3-Bisphosphoglycerate支路 ⌘ 反应关键是磷酸如何正确转移到ADP中。需要什么环境?

42
1,3-Bisphosphoglycerate
反应结构图 ADP Mg2+ 1,3-Bisphosphoglycerate PDB entry 3PGK PDB entry 1EQG

43
2,3-Bisphosphoglycerate支路
甘油酸-1,3-二 磷酸变位酶 磷酸酶

44
Reaction 8: Conversion of 3-Phosphoglycerate to 2-Phosphoglycerate
磷酸甘油变位酶 3-磷酸甘油酸 2-磷酸甘油酸 ΔG’°= 4.4 kJ/mol 碳3的磷酸转移到碳2上 ⌘ Rationale for this enzyme - repositions the phosphate to make PEP ⌘有磷酸组氨酸中间体 ⌘ 2,3-BPG 中间体产生,随即被组氨酸磷酸化

45
起始磷酸组氨酸 PDB entry 1E58

46
Reaction 9: Dehydration of 2-Phosphoglycerate to Phosphoenolpyruvate
Mg2+ 烯醇化酶 2-磷酸甘油酸 磷酸烯醇丙酮酸 ΔG’°= 1.8 kJ/mol ⌘ 烯醇化酶的作用在于促进甘油酸-2-磷酸上某些原子的重排从而形成具有较高的磷酸转移势能的高能分子。 ⌘ 氟合物能够与Mg 2+和磷酸基团形成络化物,而干扰甘油酸-2-磷酸与烯醇化的结合从而抑制该酶的活性

47
反应之前 反应之后 PDB entry 2ONE

48
Reaction 10: Transfer of Phosphoryl Group from Phosphoenolpyruvate to ADP
Mg2+,K+ 丙酮酸激酶 Phosphoenolpyruvate 丙酮酸 ΔG’°= kJ/mol ⌘ PEP转化成丙酮酸,同时产生 ATP ⌘ 产生两个ATP,可被视为糖酵解途径最后的能量回报。 ⌘ ΔG为大的负值——受到调控 激活剂?抑制剂?

49
磷酸烯醇式丙酮酸 醇式异构体 酮式异构体 丙酮酸 大家课后总结!

50
NADH的去路 糖酵解消耗NAD+,产生NADH 换个角度思考:NADH如果不及时消耗,会引起NAD+不足,导致细胞死亡 人细胞:
有氧状况下: 通过两种方式进入线粒体,进入有氧氧化产生大量ATP 无氧状况下: 在细胞质中通过乳酸脱氢酶(lactate dehydrogenase)产生NAD+,继续进入参与糖酵解 (红细胞,运动状态下的肌细胞)
产生NAD+,继续进入参与糖酵解. (红细胞,运动状态下的肌细胞)")
52
进入线粒体后进行生物氧化

53
丙酮酸的去路 脂肪酸不能转变为丙酮酸 Phosphoenolpyruvate 乳酸发酵 乳酸 丙酮酸 乙醛 乙醇 Alanine 乙醇发酵
ATP+GTP ADP+GDP+Pi 乳酸发酵 ADP NADH NAD+ ATP 乳酸 CO2 NADH NAD+ Amino Acids α-keto 丙酮酸 乙醛 乙醇 Alanine ATP+HCO3- NAD++CoA NADH 乙醇发酵 ADP+Pi Citrate 脂肪酸 Fatting Acids TCA cycle Oxidative Phosphorylation Oxaloacetate CO2 +H2O 乙酰辅酶A

54
请大家课后总结! 课前预习糖代谢剩下部分。 谢谢大家!



 反应部位 器官,细胞内定位 生理意义 代谢调节 主要调节点,主要变构抑制剂、变构激活剂 各代谢途径之间的联系和调控.>")


 合成代谢------需要能量 物质代谢 能量代谢>")









 電子傳遞鏈 傳導概論>")





