Download presentation
1
第25章 脂类代谢 一 脂类的酶促降解 二 脂肪的分解代谢 三 脂肪的合成代谢 四 磷脂的代谢 五 胆固醇的代谢

2
脂类主要包括甘油三酯(脂肪)、磷脂和类固醇等。脂类代谢是指在生物细胞内上述各类物质的生物合成和分解过程。脂类代谢对于生命活动具有重要意义。
(1)脂肪在动物体内和植物种子及果实中大量存储。脂肪在氧化时可以比其他能源物质提供更多的能量。每克脂肪氧化时可释放出38.9 kJ 的能量,每克糖和蛋白质氧化时释放的能量仅分别为17.2 kJ和23.4 kJ。 (2)许多类脂及其衍生物具有重要生理作用。脂类代谢的中间产物是合成激素、胆酸和维生素等的基本原料,对维持机体的正常活动有重要影响作用。 (3)人类的某些疾病如动脉粥样硬化、脂肪肝和酮尿症等都与脂类代谢紊乱有关。
脂肪在动物体内和植物种子及果实中大量存储。脂肪在氧化时可以比其他能源物质提供更多的能量。每克脂肪氧化时可释放出38.9 kJ 的能量,每克糖和蛋白质氧化时释放的能量仅分别为17.2 kJ和23.4 kJ。 (2)许多类脂及其衍生物具有重要生理作用。脂类代谢的中间产物是合成激素、胆酸和维生素等的基本原料,对维持机体的正常活动有重要影响作用。 (3)人类的某些疾病如动脉粥样硬化、脂肪肝和酮尿症等都与脂类代谢紊乱有关。")
3
脂肪动员: 贮存于脂肪细胞中的甘油三酯在激素敏感脂肪酶(hormone sensitive tri-glyceride lipase, HSL)的催化下水解并释放出脂肪酸,供给全身各组织细胞摄取利用的过程称为脂肪动员。
的催化下水解并释放出脂肪酸,供给全身各组织细胞摄取利用的过程称为脂肪动员。")
4
一 脂类的酶促降解

6
磷脂酶A1(B1) CH2OCOR1 CHOCOR2 CH2O—P—O—X 磷脂酶A2(B2) 磷脂酶D 磷脂酶C
 CH2OCOR1 CHOCOR2 CH2O—P—O—X 磷脂酶A2(B2) 磷脂酶D 磷脂酶C")
7
二 脂肪的分解代谢 (一)甘油的氧化
甘油的氧化")
8
甘油经血液输送到肝脏后,在ATP存在下,由甘油激酶催化,转变成-磷酸甘油。这是一个不可逆反应过程。
-磷酸甘油在脱氢酶(含辅酶NAD+)作用下,脱氢形成磷酸二羟丙酮。 在哪里进行?
作用下,脱氢形成磷酸二羟丙酮。 在哪里进行?")
9
磷酸二羟丙酮是糖酵解途径的一个中间产物,它可以沿着糖酵解途径的逆过程合成葡萄糖及糖原;也可以沿着糖酵解正常途径形成丙酮酸,再进入三羧酸循环被完全氧化。

10
(二)脂肪酸的β-氧化 脂肪酸的-氧化在线粒体基质中进行
脂肪酸的β-氧化 脂肪酸的-氧化在线粒体基质中进行")
11
脂肪酸的-氧化作用是指脂肪酸在氧化分解时,碳链的断裂发生在脂肪酸的-位,即脂肪酸碳链的断裂方式是每次切除2个碳原子。
脂肪酸的-氧化是含偶数碳原子或奇数碳原子饱和脂肪酸的主要分解方式。

12
1、脂肪酸的活化 脂肪酸进入细胞后,首先在线粒体外或胞浆中被活化,形成脂酰CoA,然后进入线粒体进行氧化。
在脂酰CoA合成酶(外膜)催化下,由ATP提供能量,将脂肪酸转变成脂酰CoA: 两个高能键,总体不可逆
催化下,由ATP提供能量,将脂肪酸转变成脂酰CoA: 两个高能键,总体不可逆.")
13
在线粒体外膜或内质网进行此反应过程。 R-COOH + HSCoA+ ATP R-CO~SCoA + AMP + PPi 脂肪酸硫激酶

14
HOOC-CH2-CH(OH)-CH2-N+-(CH3)3
2、进入 在线粒体外生成的脂酰CoA需进入线粒体基质才能被氧化分解,此过程必须要由肉碱(肉毒碱)携带,借助于两种肉碱脂肪酰转移酶(酶Ⅰ和酶Ⅱ)催化的移换反应才能完成。其中肉碱脂肪酰转移酶Ⅰ是脂肪酸β-氧化的关键酶。 HOOC-CH2-CH(OH)-CH2-N+-(CH3)3 3-羟基-4-三甲氨基丁酸
携带,借助于两种肉碱脂肪酰转移酶(酶Ⅰ和酶Ⅱ)催化的移换反应才能完成。其中肉碱脂肪酰转移酶Ⅰ是脂肪酸β-氧化的关键酶。 HOOC-CH2-CH(OH)-CH2-N+-(CH3)3. 3-羟基-4-三甲氨基丁酸.")
15
脂酰CoA转运入线粒体 长于10个碳原子的脂酰CoA,不能自由穿梭,需要借助一种特殊的载体肉毒碱(3-羟基-4-三甲氨基丁酸)才能转运到线粒体内。 脂酰CoA在线粒体膜间腔中的肉毒碱脂酰转移酶I催化下,与肉毒碱反应,生长脂酰肉毒碱。 脂酰肉毒碱在线粒体内膜的移位酶帮助下穿过内膜,并与线粒体基质中的CoA在酶II的作用下,重新生成脂酰CoA, 释放出肉毒碱。 肉毒碱再在移位酶帮助下,回到线粒体的膜间腔和细胞质中。

17
脂酰CoA进入线粒体的过程 * 肉碱脂肪酰转移酶Ⅰ在哪里? 线粒体的外膜、膜间隙、内膜外侧 移位酶的两个作用:脂酰肉碱进基质 肉碱回膜间隙
肉碱脂肪酰转移酶Ⅰ的作用:?

18
3、-氧化的反应过程 脂酰CoA在线粒体的基质中进行氧化分解。
反应产物是比原来的脂酰CoA减少了2个碳的新的脂酰CoA。 如此反复进行,直至脂酰CoA全部变成乙酰CoA。

19
β-氧化过程由四个连续的酶促反应组成: ① 脱氢 ② 水化 ③ 再脱氢 ④ 硫解

20
β-氧化的反应过程 脱氢 脂酰CoA脱氢酶 水化酶 水化 硫解 硫解酶 β-羟脂酰CoA脱氢酶 再脱氢

21
3、-氧化:(1)脱氢 脂酰CoA在脂酰CoA脱氢酶的催化下,在-和-碳原子上各脱去一个氢原子,生成反式,-烯脂酰CoA,氢受体是FAD。
脱氢 脂酰CoA在脂酰CoA脱氢酶的催化下,在-和-碳原子上各脱去一个氢原子,生成反式,-烯脂酰CoA,氢受体是FAD。")
22
3、-氧化:(2)水化 在烯脂酰CoA水合酶催化下,,-烯脂酰CoA水化,生成L(+)--羟脂酰CoA
水化 在烯脂酰CoA水合酶催化下,,-烯脂酰CoA水化,生成L(+)--羟脂酰CoA")
23
3、-氧化: (3)再脱氢 -羟脂酰CoA在脱氢酶催化下,脱氢生成-酮脂酰CoA。反应的氢受体为NAD+。此脱氢酶具有立体专一性,只催化L(+)--羟脂酰CoA的脱氢。
再脱氢 -羟脂酰CoA在脱氢酶催化下,脱氢生成-酮脂酰CoA。反应的氢受体为NAD+。此脱氢酶具有立体专一性,只催化L(+)--羟脂酰CoA的脱氢。")
24
3、-氧化: (4)硫解 在-酮脂酰CoA硫解酶催化下,-酮脂酰CoA与CoA作用,生成1分子乙酰CoA和1分子比原来少两个碳原子的脂酰CoA。少了两个碳原子的脂酰CoA ,可以重复上述反应过程,一直到完全分解成乙酰CoA。脂肪酸通过-氧化生成的乙酰CoA,一部分用来合成新的脂肪酸和其它生物分子,大部分则进入三羧酸循环完全氧化。

25
脂肪酸-氧化产生的能量 脂肪酸的完全氧化可以产生大量的能量。例如软脂酸(含16碳)经过7次-氧化,可以生成8个乙酰CoA,每一次-氧化,还将生成1分子FADH2和1分子NADH。软脂酸完全氧化的反应式为: C16H31CO-SCoA + 7 CoA-SH + 7 FAD + NAD+ +7 H2O 8 CH3CO-SCoA + 7 FADH2 + 7 NADH + 7 H+

26
脂肪酸-氧化产生的能量 按照一个NADH产生2.5个ATP,1个FADH2产生1.5个ATP, 1个乙酰CoA完全氧化产生10个ATP计算,1分子软脂酰CoA在分解代谢过程中共产生108个ATP。 由于软脂酸转化成软脂酰CoA时消耗了1分子ATP中的两个高能磷酸键的能量(ATP分解为AMP, 可视为消耗了2个ATP),因此,1分子软脂酸完全氧化净生成 108 – 2 = 106个ATP 效率比葡萄糖高
,因此,1分子软脂酸完全氧化净生成 108 – 2 = 106个ATP. 效率比葡萄糖高.")
27
脂肪酸的β-氧化

28
脂肪酸β氧化的五个步骤: 1.脂肪酸活化(脂酰CoA合酶),外膜,ATP-AMP 利用肉碱进行穿梭进入基质
2.氧化(脂酰CoA脱氢酶)——烯酰CoA,FADH2 3.水化(烯酰CoA水合酶)——3-羟脂酰CoA 4.氧化( 3-羟脂酰CoA脱氢酶)——3-酮脂酰CoA,NADH 5.断裂(β酮硫解酶)+CoA——乙酰CoA+少两个碳原子的脂酰CoA 再循环,所以软脂酸经7轮氧化,得8个乙酰COA
——烯酰CoA,FADH2. 3.水化(烯酰CoA水合酶)——3-羟脂酰CoA. 4.氧化( 3-羟脂酰CoA脱氢酶)——3-酮脂酰CoA,NADH. 5.断裂(β酮硫解酶)+CoA——乙酰CoA+少两个碳原子的脂酰CoA. 再循环,所以软脂酸经7轮氧化,得8个乙酰COA.")
29
不饱和脂肪酸的β氧化 与饱和脂肪酸的前三轮相同
当遇到双键时,由烯酰CoA异构酶将顺式双键异构为反式双键,不需脱氢酶作用,所以少生成1 分子FADH2,即1.5ATP

30
脂肪酸β-氧化的特点: ① β-氧化过程在线粒体基质内进行; ② β-氧化为一循环反应过程,由脂肪酸氧化酶系催化,反应不可逆;
③ 需要FAD,NAD,CoA为辅助因子; ④ 每循环一次,生成一分子FADH2,一分子NADH,一分子乙酰CoA和一分子减少两个碳原子的脂酰CoA。

31
(三)脂肪酸氧化的其他途径 1. 奇数碳链脂肪酸的氧化 17碳的脂肪酸氧化成7乙酰CoA和1个丙酰COA
1. 奇数碳链脂肪酸的氧化 17碳的脂肪酸氧化成7乙酰CoA和1个丙酰COA 丙酰CoA经过羧化、异构形成琥珀酰CoA进入柠檬酸循环(P404,P242) 所以生物素、B12等 主要存在于反胄动物体内 2.α氧化: 3.ω氧化
 所以生物素、B12等. 主要存在于反胄动物体内. 2.α氧化: 3.ω氧化.")
32
2、-氧化:在植物种子萌发时,脂肪酸的-碳被氧化成羟基,生成-羟基酸。-羟基酸可进一步脱羧、氧化转变成少一个碳原子的脂肪酸。上述反应由单氧化酶催化,需要有O2、Fe2+和抗坏血酸等参加。
α-氧化 RCH2COOH RCH(OH)COOH RCOCOOH RCOOH + CO2
COOH. RCOCOOH. RCOOH + CO2.")
33
CH3(CH2)nCOO- HOCH2(CH2)nCOO- -OOC(CH2)nCOO- 3.-氧化:在动物体中,C10 或C11脂肪酸的碳链末端碳原子(-碳原子)可以先被氧化,形成二羧酸。二羧酸进入线粒体内后,可以从分子的任何一端进行-氧化,最后生成的琥珀酰CoA可直接进入三羧酸循环。
nCOO- HOCH2(CH2)nCOO- -OOC(CH2)nCOO- 3.-氧化:在动物体中,C10 或C11脂肪酸的碳链末端碳原子(-碳原子)可以先被氧化,形成二羧酸。二羧酸进入线粒体内后,可以从分子的任何一端进行-氧化,最后生成的琥珀酰CoA可直接进入三羧酸循环。")
34
乙酰CoA的结局 TCA循环 合成胆固醇 合成脂肪酸 当还有过多的乙酰CoA时会在肝脏中生成酮体 ——乙酰乙酸,D-β羟丁酸和丙酮

35
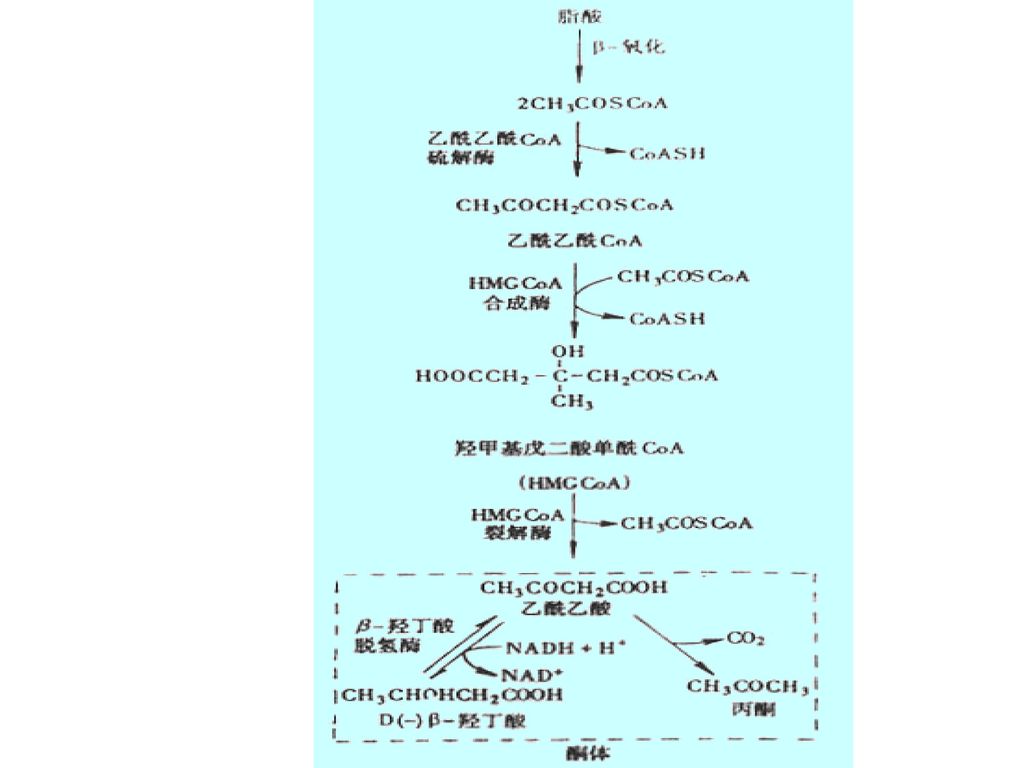
(四) 酮体的生成及利用: 脂肪酸在肝脏中氧化分解所生成的乙酰乙酸、β-羟丁酸和丙酮三种中间代谢产物,统称为酮体。
CH3COCH2COOH 乙酰乙酸 CH3CH(OH)CH2COOH β-羟丁酸 CH3COCH 丙酮
CH2COOH β-羟丁酸. CH3COCH3 丙酮.")
36
酮体的生成和利用 1. 酮体的生成(肝组织)
")
37
有些组织对酮体的利用甚至优于葡萄糖的利用,如心脏及饥饿时的大脑
脂肪组织 脂肪酸 乳腺 脑 肝脏 酮体 肾脏 肌肉 饥饿时酮体的利用 肠道

39
1.酮体的生成: 酮体主要在肝脏的线粒体中生成,其合成原料为乙酰CoA。
(1) 两分子乙酰CoA在乙酰乙酰CoA硫解酶的催化下,缩合生成一分子乙酰乙酰CoA。 2 CH3CO-CoA CH3COCH2CO-CoA + HSCoA 乙酰乙酰硫解酶
 两分子乙酰CoA在乙酰乙酰CoA硫解酶的催化下,缩合生成一分子乙酰乙酰CoA。 2 CH3CO-CoA CH3COCH2CO-CoA + HSCoA. 乙酰乙酰硫解酶.")
40
* (2) 乙酰乙酰CoA再与1分子乙酰CoA缩合,生成HMG-CoA。HMG-CoA合成酶是酮体生成的关键酶。
CH3COCH2CO-CoA + CH3CO-CoA HOOCCH2C(OH)(CH3)CH2CO-SCoA + HSCoA * HMG-CoA合成酶 HMG:β-羟基-β-甲基戊二酸
(CH3)CH2CO-SCoA + HSCoA. * HMG-CoA合成酶. HMG:β-羟基-β-甲基戊二酸.")
41
(3) HMG-CoA裂解生成1分子乙酰乙酸和1分子乙酰CoA。
HOOCCH2C(OH)(CH3)CH2CO-SCoA CH3COCH2COOH + CH3CO-CoA HMG-CoA裂解酶
(CH3)CH2CO-SCoA. CH3COCH2COOH + CH3CO-CoA. HMG-CoA裂解酶.")
42
(4) 乙酰乙酸在β-羟丁酸脱氢酶的催化下,加氢还原为β-羟丁酸。
CH3COCH2COOH + NADH + H+ CH3CH(OH)CH2COOH + NAD+ (5) 乙酰乙酸也可自发脱羧生成丙酮。 CH3COCH2COOH CH3COCH3 + CO2 β-羟丁酸脱氢酶
CH2COOH + NAD+ (5) 乙酰乙酸也可自发脱羧生成丙酮。 CH3COCH2COOH CH3COCH3 + CO2. β-羟丁酸脱氢酶.")
44
2. 酮体的氧化(肝外组织)(了解) CH3CHOHCH2COOH CH3COCH2COOH
2. 酮体的氧化(肝外组织)(了解) CH3CHOHCH2COOH CH3COCH2COOH CH3COCH2COOH + -OOCCH2CH2COSCoA 琥珀酰辅酶A转硫酶 CH3COCH2COSCoA + -OOCCH2CH2COO- CH3COCH2COOH+ CoASH +ATP 乙酰乙酸硫激酶 CH3COCH2COSCoA + AMP + PPi CH3COCH2COSCoA + CoASH → 2CH3COSCoA
(了解) CH3CHOHCH2COOH. CH3COCH2COOH. CH3COCH2COOH + -OOCCH2CH2COSCoA. 琥珀酰辅酶A转硫酶. CH3COCH2COSCoA + -OOCCH2CH2COO- CH3COCH2COOH+ CoASH +ATP. 乙酰乙酸硫激酶. CH3COCH2COSCoA + AMP + PPi. CH3COCH2COSCoA + CoASH → 2CH3COSCoA.")
45
(1) β-羟丁酸在β-羟丁酸脱氢酶的催化下脱氢,生成乙酰乙酸。
了解 酮体利用的基本过程为: (1) β-羟丁酸在β-羟丁酸脱氢酶的催化下脱氢,生成乙酰乙酸。 CH3CH(OH)CH2COOH + NAD+ CH3COCH2COOH + NADH + H+ β-羟丁酸脱氢酶
 β-羟丁酸在β-羟丁酸脱氢酶的催化下脱氢,生成乙酰乙酸。 CH3CH(OH)CH2COOH + NAD+ CH3COCH2COOH + NADH + H+ β-羟丁酸脱氢酶.")
46
(2) 乙酰乙酸在琥珀酰CoA转硫酶或乙酰乙酸硫激酶的催化下转变为乙酰乙酰CoA。
了解 (2) 乙酰乙酸在琥珀酰CoA转硫酶或乙酰乙酸硫激酶的催化下转变为乙酰乙酰CoA。 CH3COCH2COOH + HOOCCH2CH2CO-CoA CH3COCH2CO-CoA + HOOCCH2CH2COOH 琥珀酰CoA转硫酶
 乙酰乙酸在琥珀酰CoA转硫酶或乙酰乙酸硫激酶的催化下转变为乙酰乙酰CoA。 CH3COCH2COOH + HOOCCH2CH2CO-CoA. CH3COCH2CO-CoA + HOOCCH2CH2COOH. 琥珀酰CoA转硫酶.")
47
(3) 乙酰乙酰CoA在乙酰乙酰CoA硫解酶的催化下,裂解为两分子乙酰CoA。
了解 (3) 乙酰乙酰CoA在乙酰乙酰CoA硫解酶的催化下,裂解为两分子乙酰CoA。 CH3COCH2CO-CoA + HSCoA 2 CH3CO-CoA (4) 生成的乙酰CoA进入三羧酸循环彻底氧化分解 乙酰乙酰CoA硫解酶
 乙酰乙酰CoA在乙酰乙酰CoA硫解酶的催化下,裂解为两分子乙酰CoA。 CH3COCH2CO-CoA + HSCoA. 2 CH3CO-CoA. (4) 生成的乙酰CoA进入三羧酸循环彻底氧化分解. 乙酰乙酰CoA硫解酶.")
48
了解 心、肾、脑细胞 心、肾、脑、骨骼肌细胞

49
丙酮去路 (1)随尿排出 (2)直接从肺部呼出 (3)转变为丙酮酸或甲酰基及乙酰基 (4)中毒、毒性 饥饿 草酰乙酸合成糖 乙酰CoA过多
输出肝脏供能 合成酮体

50
酮体生成及利用的生理意义: (1) 在正常情况下,酮体是肝脏输出能源的一种形式;为什么不异生?
(2) 在饥饿或疾病情况下,为心、脑等很多重要器官提供必要的能源。 (3)不能被肝脏和红细胞利用。肝脏没有利用酮体的酶,红细胞没有线粒体
 在饥饿或疾病情况下,为心、脑等很多重要器官提供必要的能源。 (3)不能被肝脏和红细胞利用。肝脏没有利用酮体的酶,红细胞没有线粒体.")
51
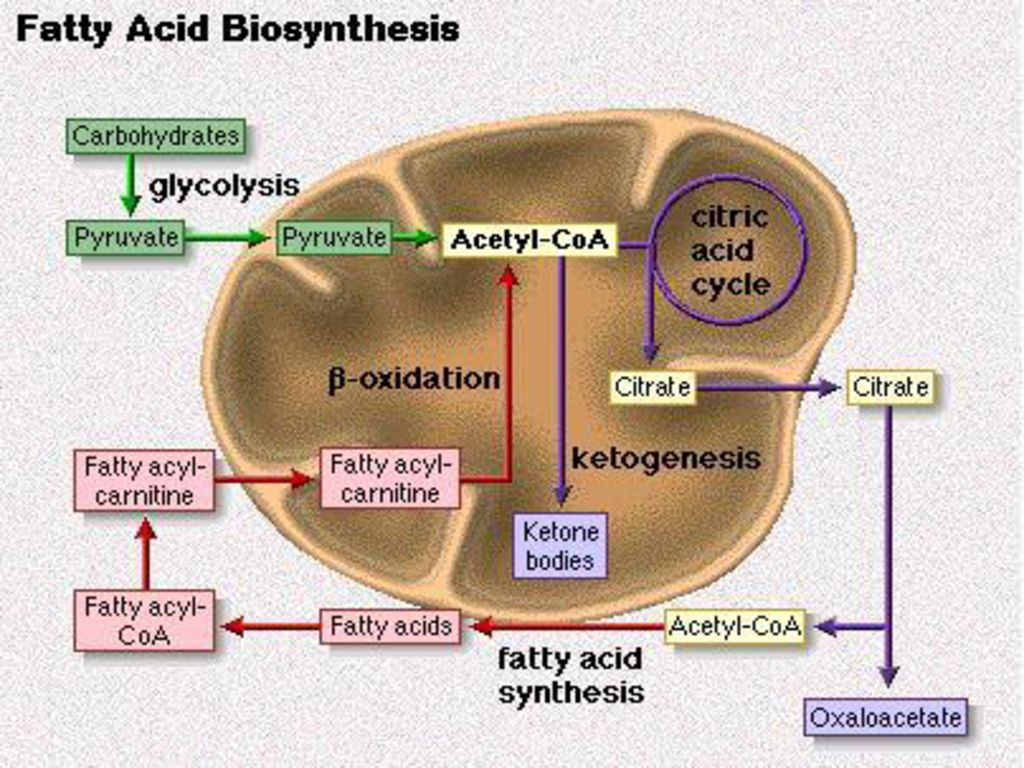
脂肪的合成代谢 脂肪酸分解在线粒体基质中 合成在细胞溶胶中 脂肪动员:储存的脂肪释放出游离的脂肪酸和甘油,并将脂肪酸运到肝脏。
其中,脂肪酸在线粒体的基质中分解 甘油在细胞溶胶中分解

53
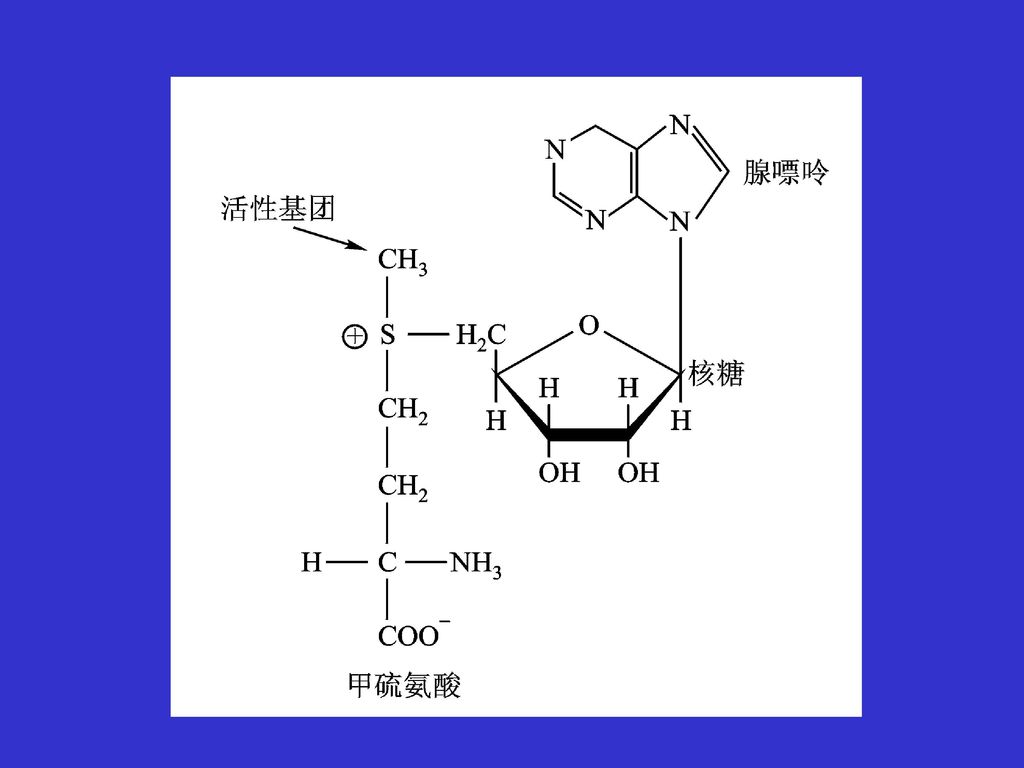
过度的脂肪动员会产生脂肪肝 脂类和糖原的代谢异常 膳食中的胆碱和甲硫氨酸(卵磷脂)会抑制脂肪肝的形成,抗脂肪肝剂、脂蛋白(运输脂质)
胆碱的合成离不开丝氨酸与SAM,丝氨酸提供碳骨架;S-腺酐甲硫氨酸(SAM)提供甲基 过度的脂肪动员导致脂蛋白的减少,影响脂从肝脏的运输
提供甲基. 过度的脂肪动员导致脂蛋白的减少,影响脂从肝脏的运输.")
55
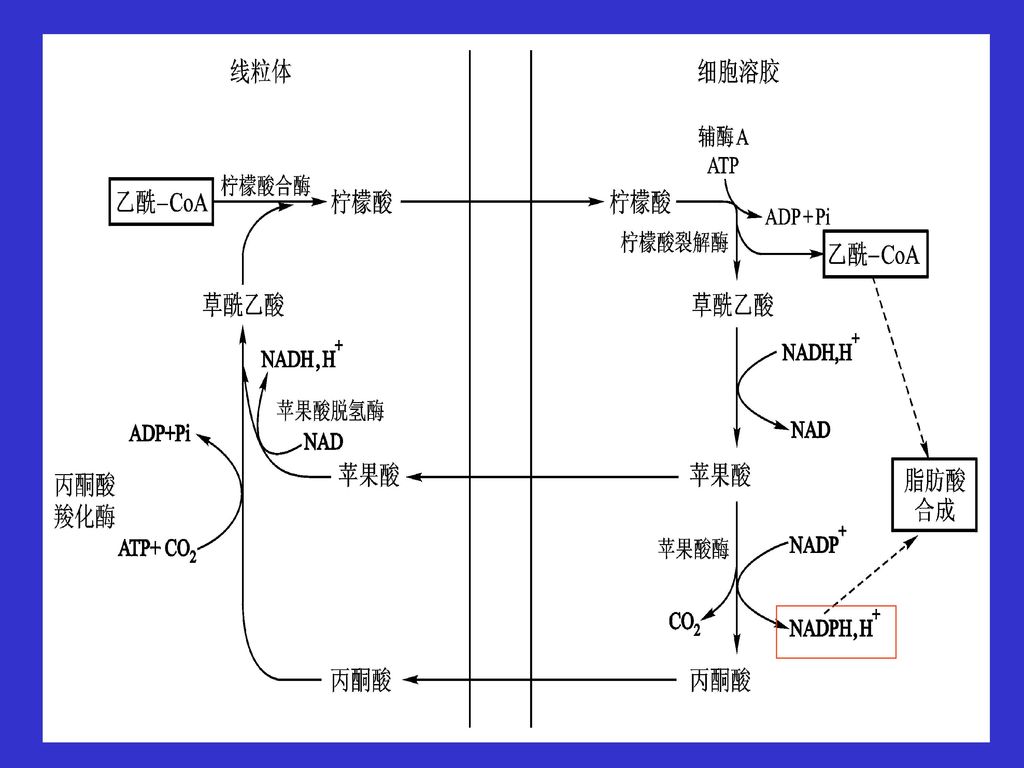
一、脂肪酸的生物合成的准备 乙酰CoA,丙二酸单酰CoA,脂肪酸合酶 1、乙酰-CoA的准备: 是合成底物 但是乙酰-CoA产生于线粒体
脂肪酸的合成在细胞溶胶 必须经过三羧酸转运体系 通过柠檬酸离开线粒体到细胞溶胶 通过苹果酸或丙酮酸进入线粒体

58
若是通过丙酮酸穿梭则 每输出一分子的乙酰CoA形成1分子NADPH 用于脂肪酸的合成 经柠檬酸-丙酮酸穿梭作用将线粒体内生成的乙酰CoA运至胞液

59
2.丙二酸单酰CoA的合成准备: 在乙酰CoA羧化酶的催化下,将乙酰CoA羧化为丙二酸单酰CoA。乙酰CoA羧化酶受柠檬酸和异柠檬酸的变构激活,受长链脂酰CoA的变构抑制 CH3CO-SCoA + HCO3- + H+ + ATP HOOC-CH2-CO-SCoA + ADP + Pi * 乙酰CoA羧化酶 (生物素)
")
60
羧化 转羧 总反应 乙酰CoA在乙酰CoA羧化酶形成丙二酸单酰CoA
三成分:1.BCCP,生物素羧基载体蛋白,生物素的载体 生物素羧化酶 转羧酶 羧化 转羧 总反应 耗ATP

64
3、脂肪酸合酶复合体的准备 合成反应由脂肪酸合成酶系催化。
脂肪酸合成时碳链的缩合延长过程是一循环反应过程。每经过一次循环反应,延长两个碳原子。

65
在低等生物中,脂肪酸合成酶系是一种由1分子脂酰基载体蛋白(ACP)和7种酶单体所构成的多酶复合体
和7种酶单体所构成的多酶复合体")
66
动物中,7种酶和酰基载体蛋白(ACP),单一的多功能多肽链,有利于不同酶的协作
也是CoA的活性基团 ACP在合成中的作用相当于CoA在分解在中的作用

67
磷酸基团连接蛋白,巯基连接脂酰基,起转移脂酰基的作用

68
原核生物脂肪酸合成酶系

69
真核生物脂肪酸合成酶系

70
二、脂肪酸合成的步骤 7个酶7个步骤 1.启动 乙酰CoA-ACP 转酰酶 2.装载 丙二酰CoA-ACP 转酰酶

72
1.启动 乙酰CoA-ACP 转酰酶 2.装载 丙二酰CoA-ACP 转酰酶 3.缩合 β酮酰ACP合酶

73
4.还原 β酮酰ACP还原酶 5.脱水 β羟酰ACP脱水酶

74
6.还原 烯酰ACP还原酶 7.释放 软脂酰ACP硫脂酶 与下一个丙二酸单酰ACP作用,再延长两个碳原子,一直到16个 软脂酰ACP+H2O——软脂酸+HS-ACP

75
从头合成

77
总反应: 7个ATP是生成丙二酸单酰CoA时需提供 14个NADPH中8个可来自于乙酰CoA的穿梭 另外6个可来自于?

78
三、脂肪酸合成途径与β氧化的比较: 场所不同 载体不同 穿梭机制:合成使用三羧酸转运,分解使用肉碱机制
都以二碳单位进行代谢,合成需要丙二酸单酰CoA 合成是甲基端开始,分解是羧基端开始 合成需要NADPH,分解需要NAD与FAD 动物体脂肪酸合成代谢的酶在一条多肽链上 以16碳为例,都需经过7个循环

79
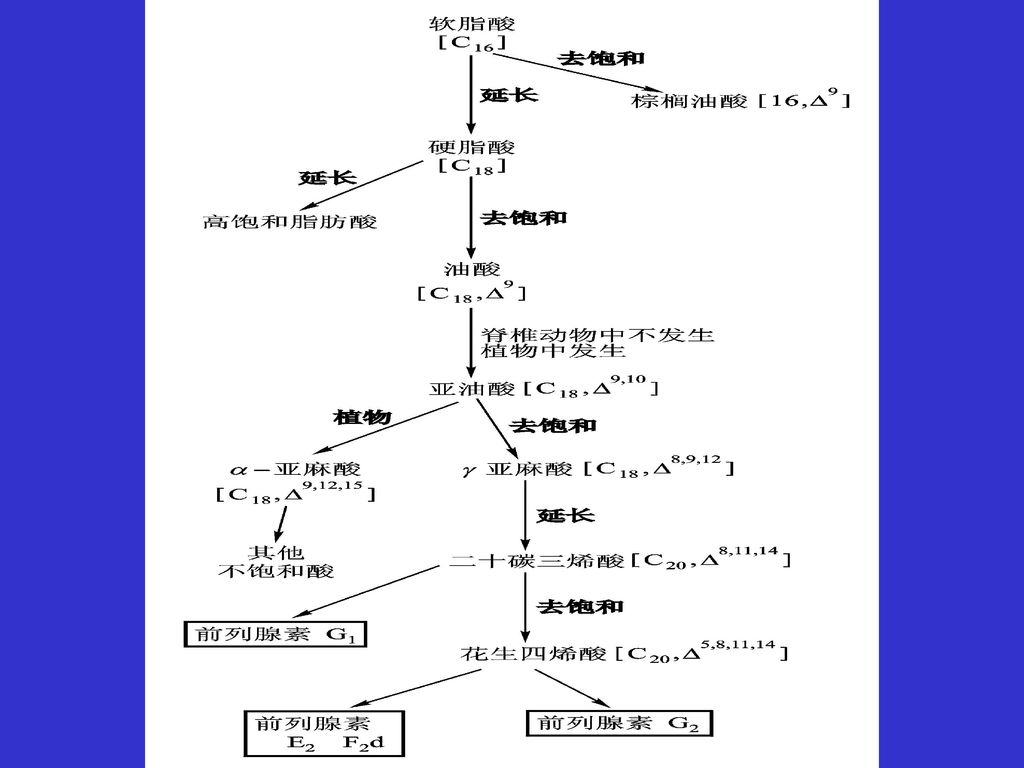
脂肪酸进一步的延长与去饱和 都需要以软脂酸为前体 延长:不同部位延长机制有差异 线粒体:降解的逆反应,NADPH代替FAD
内质网:NADPH代替FAD,用CoA而不是ACP 去饱和:脂酰CoA去饱和酶在第9位碳原子上引入双键 动物缺乏在其他位置引入双键能力 故亚油酸和亚麻酸是必须脂肪酸

80
线粒体系统(基本是β-氧化的逆过程)
")
81
必需脂肪酸: 在饱和脂肪酸中引入双键(加氧酶) (1)亚油酸(Δ9,12—十八二烯酸) (2)亚麻酸(Δ9,12,15—十八三烯酸)
(3)花生四烯酸(Δ5,8,11,12—二十四烯酸) 在饱和脂肪酸中引入双键(加氧酶) CH3(CH2)14COSCoA + NADPH + H+ CH3(CH2)5CH=CH(CH2)7COSCoA + NADP+ + 2H2O
花生四烯酸(Δ5,8,11,12—二十四烯酸) 在饱和脂肪酸中引入双键(加氧酶) CH3(CH2)14COSCoA + NADPH + H+ CH3(CH2)5CH=CH(CH2)7COSCoA + NADP+ + 2H2O.")
83
四、脂肪酸合成的调节 合成与分解的关系 乙酰CoA羧化酶是限速酶,关键 1.软脂酰CoA反馈抑制
2.乙酰CoA增多(柠檬酸从线粒体到胞外),ATP别构激活 3.共价修饰,去磷酸化即活化(糖原的合成、脂类的合成) 合成与分解的关系 合成中产生的丙二酸单酰CoA可以抑制分解中的肉碱脂酰转移酶I,关闭分解
,ATP别构激活. 3.共价修饰,去磷酸化即活化(糖原的合成、脂类的合成) 合成与分解的关系. 合成中产生的丙二酸单酰CoA可以抑制分解中的肉碱脂酰转移酶I,关闭分解.")
85
五、脂质的代谢 1、脂肪的合成(三酰甘油) 获得磷酸甘油的两个途径: 1)甘油 + ATP → 磷酸甘油 + ADP
脂肪动员的甘油运到肝脏:甘油激酶 2)磷酸二羟丙酮 + NADH + H+ → 磷酸甘油 + NAD+ 穿梭机制,磷酸甘油脱氢酶
磷酸二羟丙酮 + NADH + H+ → 磷酸甘油 + NAD+ 穿梭机制,磷酸甘油脱氢酶.")
86
磷脂酸 CH2OCOR CHOCOR CH2O P CH2OH CHOH CH2O P 2RCOSCoA + + 2CoASH 磷脂酸磷酸酶
酰基转移酶 2RCOSCoA + + 2CoASH 磷脂酸磷酸酶 RCOSCoA CH2OCOR CHOCOR CH2OHCOR CHOHCOR CH2OH 酰基转移酶

87
2、磷脂的代谢(分解:磷脂酶) 磷脂的合成能力:肝>肠>肌肉>脑组织。 需要CTP的参与,CDP是脂质的重要载体
磷脂酰胆碱和磷脂酰乙醇胺通过类似代谢途径合成。 合成过程中所需胆碱及乙醇胺以CDP-胆碱和CDP-乙醇胺的形式提供。

88
甘油磷脂的代谢: 甘油磷脂的基本结构: CH2-O-CO-R' | R"-CO-O-CH CH2-O-PO3H-X

89
磷脂酸磷酸酶 二酰甘油 真核生物:CDP 二酰甘油或CDP胆碱 原核生物只能是CDP二脂酰甘油

90
甘油磷酯合成途径二

91
卵磷脂的合成(真核) 胆碱(不能在动物体生成)——磷酸胆碱(胆碱激酶) 磷酸胆碱+CTP——CDP胆碱(磷酸胆碱胞苷转移酶)限速酶
或由磷脂酰丝氨酸经SAM甲基化而成

92
其他甘油磷脂的合成二(同原核生物)
")
93
3、 胆固醇的代谢 (一)胆固醇的消化和吸收 1. 再循环(肠肝循环):食入的胆固醇吸收后又可通过胆汁或肠壁而排入肠腔,它们和膳食中的胆固醇混合在一起而被重新吸收。 2.吸收有限:肠道吸收胆固醇的能力极为有限。
:食入的胆固醇吸收后又可通过胆汁或肠壁而排入肠腔,它们和膳食中的胆固醇混合在一起而被重新吸收。 2.吸收有限:肠道吸收胆固醇的能力极为有限。 .")
95
(二)胆固醇的分解和转化 1、转变为胆酸。 初级胆汁酸合成的关键酶是7α-羟化酶。 2、转变成维生素 3. 转变为激素
胆固醇的分解和转化 1、转变为胆酸。 初级胆汁酸合成的关键酶是7α-羟化酶。 2、转变成维生素 3. 转变为激素")
96
转化为类固醇激素: 1.肾上腺皮质激素的合成: 2.雄激素的合成: 3.雌激素的合成: 转化为维生素D3:

97
(三)胆固醇合成的基本过程: HMG-CoA还原酶是胆固醇合成的关键酶。 1.乙酰CoA缩合生成甲羟戊酸(MVA):
此过程在胞液和微粒体进行。 2×乙酰CoA→乙酰乙酰CoA→HMG-CoA →MVA ——脱羧形成5碳的异戊二烯

98
2.甲羟戊酸缩合生成鲨烯: 6个异戊二烯——鲨烯(30碳) 3.鲨烯环化为胆固醇: 27碳
 3.鲨烯环化为胆固醇: 27碳")

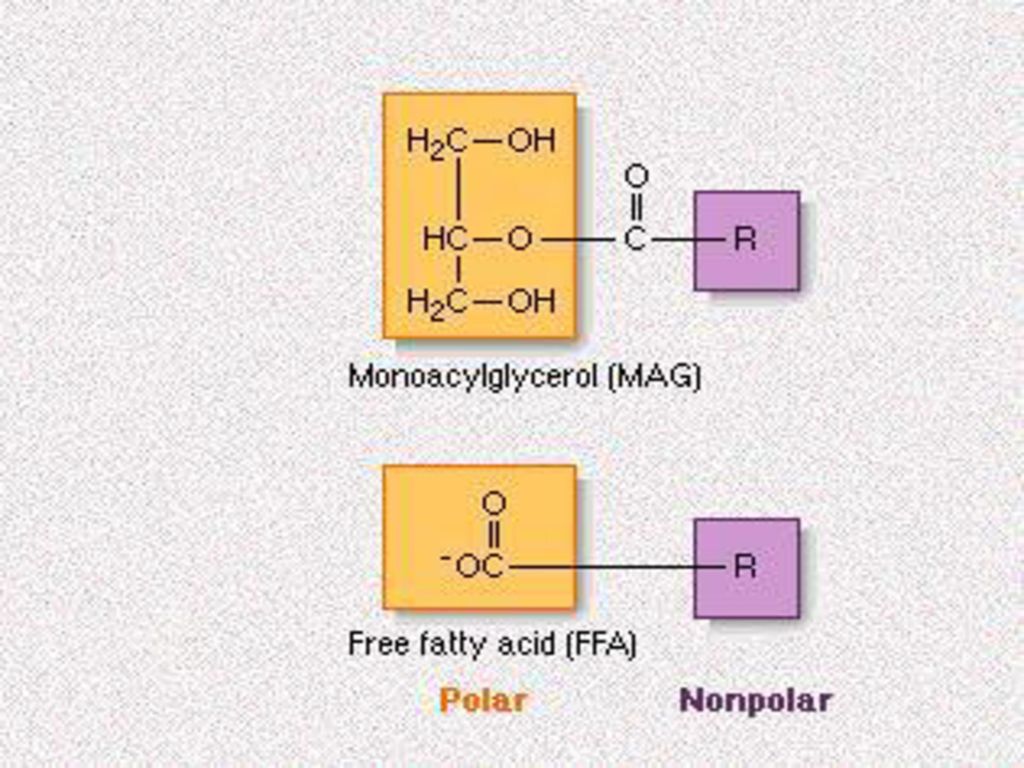










 不饱和脂肪酸的氧化 酮体 磷脂的代谢 脂肪酸代谢的调控.>")
 信号传导:激素、胞内信使.>")

 反应部位 器官,细胞内定位 生理意义 代谢调节 主要调节点,主要变构抑制剂、变构激活剂 各代谢途径之间的联系和调控.>")
 CO.>")


.>")


物质在生物体的分解及合成代谢。要求学生重点掌握脂肪酸在生物体内的氧化分解途径—β-氧化和从头合成途径,了解脂类物质的功能和其他的氧化分解途径。 思考 脂类代谢 返回.>")

>")






