Download presentation
Presentation is loading. Please wait.

1
第九章 糖 代 谢

2
本 章 内 容 糖的消化(水解)、吸收及转运。 分解代谢:糖酵解、三羧酸循环、磷酸戊糖途径、糖原分解。
合成代谢:糖异生、糖原合成、光合作用。 代谢途径的特点及意义。 糖代谢的调节。 教材第10、15章

3
第一节 糖的消化、吸收和转运 一、消化 1、α-淀粉酶(唾液淀粉酶,液化酶;胰腺)
是一种内切酶,水解-1,4-糖苷键,将淀粉随机切断成分子量较小的糊精。 2、β-淀粉酶(胰腺) 从链的非还原性末端开始,水解-1,4-糖苷键,每次切下两个葡萄糖单位——β-麦芽糖。 3、γ-淀粉酶(糖化酶) 从链的非还原性末端开始,水解-1,4-糖苷键和-1,6-糖苷键,将淀粉完全水解成葡萄糖。
 从链的非还原性末端开始,水解-1,4-糖苷键,每次切下两个葡萄糖单位——β-麦芽糖。 3、γ-淀粉酶(糖化酶) 从链的非还原性末端开始,水解-1,4-糖苷键和-1,6-糖苷键,将淀粉完全水解成葡萄糖。")
4
4、二糖酶(人及动物的小肠液和微生物中) 蔗糖酶:将蔗糖水解成D-葡萄糖和D-果糖。 乳糖酶:将乳糖水解为D-葡萄糖和D-半乳糖。 麦芽糖酶:将麦芽糖水解成D-葡萄糖。 5、纤维素酶(反刍动物肠道细菌产生) 作用于-1, 4-糖苷键,将纤维素水解成葡萄糖。 6、糖原磷酸化酶(细胞内酶) 糖原n + Pi → 糖原n 磷酸-葡萄糖
 糖原n + Pi → 糖原n 磷酸-葡萄糖.")
5
二、吸收 三、转运 小肠粘膜表面上皮细胞存在特定载体,称为Na+依赖型 葡萄糖转运体,是主动耗能的过程。 动物:以葡萄糖形式运输,称为血糖。
人:80~120mg%(4.4~6.7mmol/L)。 植物:以蔗糖形式运输。
。 植物:以蔗糖形式运输。")
6
食物 肝糖原分解 糖异生 氧化供能 肝、肌糖原 脂肪(肝、脂肪组织) 含糖物质和非糖物质 (核糖、氨基糖)(非必需AA) 血 糖
 含糖物质和非糖物质 (核糖、氨基糖)(非必需AA) 血 糖")
7
第二节 糖的分解代谢 葡萄糖进入细胞后,在一系列酶的催化下,发生分解代谢过程:
(1)糖酵解:葡萄糖 丙酮酸 乳酸。此反应过程一般在无氧条件下进行,又称为无氧分解。 (2)三羧酸循环(柠檬酸循环):丙酮酸 CO2 + H2O 。由于分子氧是此系列反应的最终受氢体,所以又称为有氧分解。 (3)其他代谢途径:磷酸己糖支路,乙醛酸循环等。
糖酵解:葡萄糖 丙酮酸 乳酸。此反应过程一般在无氧条件下进行,又称为无氧分解。 (2)三羧酸循环(柠檬酸循环):丙酮酸 CO2 + H2O 。由于分子氧是此系列反应的最终受氢体,所以又称为有氧分解。 (3)其他代谢途径:磷酸己糖支路,乙醛酸循环等。")
8
1葡萄糖 2磷酸甘油醛 需ATP 两个阶段 2磷酸甘油醛 2丙酮酸 产生ATP
一、糖酵解(glycolysis) 定义:是糖的无氧氧化,葡萄糖在缺氧或氧气供应不足的条件下进行分解生成丙酮酸且还原生成乳酸,同时放出少量能量的过程,简称EMP途径(Embden-Meyerhof pathway)。 (一) 反应过程:10步 1葡萄糖 2磷酸甘油醛 需ATP 两个阶段 磷酸甘油醛 2丙酮酸 产生ATP
 定义:是糖的无氧氧化,葡萄糖在缺氧或氧气供应不足的条件下进行分解生成丙酮酸且还原生成乳酸,同时放出少量能量的过程,简称EMP途径(Embden-Meyerhof pathway)。 (一) 反应过程:10步. 1葡萄糖 2磷酸甘油醛 需ATP 两个阶段 2磷酸甘油醛 2丙酮酸 产生ATP.")
9
1、第一阶段: G 6-P- G(关键反应) 部 位 专一性 G 的亲和力 己糖激酶 各组织 低 高 葡萄糖激酶 肝 高 低
 部 位 专一性 G 的亲和力 己糖激酶 各组织 低 高 葡萄糖激酶 肝 高 低.")
10
②6-P-G 6-P-F

11
③6-P –F 1,6 - 2P – F(限速反应)
")
12
磷酸二羟丙酮 ④ 1,6 - 2P - F 3-磷酸甘油醛

13
酵解中唯一的脱氢反应,同时与磷酸化反应相耦联(底物水平磷酸化)。
2、第二阶段: ① 第1个ATP的生成: 酵解中唯一的脱氢反应,同时与磷酸化反应相耦联(底物水平磷酸化)。 NO 016
。 NO 016.")
14
②.第2个ATP的生成: 限速反应,底物水平磷酸化

15
糖酵解总过程(P338图10-2)
")
16
糖酵解的总反应式: C6H12O6+2ADP+2H3PO4+2NAD+2C3H4O3+2ATP+2NADH+2H++2H2O
葡萄糖 丙酮酸 2C3H4O3 + 2NADH + 2H+→2C3H6O3 + 2NAD+ 丙酮酸 乳酸

17
(二) 小 结 1、部位:胞液 2、产能:2ATP 3、限速酶:己糖激酶、磷酸果糖激酶-1、丙酮酸激酶 4、意义: ◆ 酵解是糖分解代谢的共同中心途径。 ◆ 迅速提供能量,肌肉收缩。 ◆ 成熟的红细胞完全依赖酵解供能。 ◆ 丙酮酸的去路: 进入三羧酸循环(有氧条件) 乳酸发酵(无氧条件) 乙醇发酵(无氧条件,微生物,丙酮酸脱羧酶)
 乳酸发酵(无氧条件) 乙醇发酵(无氧条件,微生物,丙酮酸脱羧酶)")
18
COOH C=O CH3 + NADH + H+ 乳酸脱氢酶 CHOH + NAD+ 丙酮酸 乳酸 COOH C=O CH3 丙酮酸脱羧酶 CHO + CO2 CH2OH + NADH + H+ 乙醇脱氢酶 + NAD+ 丙酮酸 乙醛 TPP,Mg2+ 乙醇

19
糖原及其他单糖进入酵解的途径 膳食中糖类 胞内贮存多糖 糖原 ↓ G-1-P ↑↓ G-6-P F-6-P 磷酸二羟丙酮 3-磷酸甘油醛
丙酮酸 半乳糖 葡萄糖 果糖 ↓肝 F-1-P 乳糖 麦芽糖 蔗糖 甘露糖→甘露糖-6-P 甘油醛 肌肉、肾

20
G-6-P - 己糖激酶 葡萄糖激酶 hexokinase glucokinase (三) 糖酵解的调节(p337)
通过各种变构剂对三个关键酶进行变构调节 1. 己糖激酶或葡萄糖激酶: 葡萄糖激酶是肝脏调节葡萄糖吸收的主要关键酶。 己糖激酶 hexokinase 葡萄糖激酶 glucokinase G-6-P 长链脂酰CoA 乙酰CoA -

21
6-phosphofructokinase-1
2. 6-磷酸果糖激酶-1(PFK1): 是调节糖酵解代谢途径流量的主要因素。 ATP 柠檬酸 H+ ADP、AMP、Pi 2,6-二磷酸果糖 - + 6-磷酸果糖激酶-1 6-phosphofructokinase-1
: 是调节糖酵解代谢途径流量的主要因素。 ATP. 柠檬酸. H+ ADP、AMP、Pi. 2,6-二磷酸果糖 磷酸果糖激酶-1. 6-phosphofructokinase-1.")
22
3. 丙酮酸激酶: 控制酵解途径代谢物的输出。 丙酮酸激酶 pyruvate kinase ATP 丙氨酸(肝) 1,6-二磷酸果糖 - +
 1,6-二磷酸果糖 - +")
24
二、糖的有氧氧化 定义:葡萄糖通过糖酵解产生的丙酮酸,在有氧条件下的氧化分解,最终产生H2O和CO2,并释放大量能量的过程,是糖氧化的主要方式。 第一阶段:丙酮酸的氧化脱羧(丙酮酸乙酰辅酶A) 第二阶段:三羧酸循环(tricarboxylic acid cycle,TCA cycle,又称柠檬酸循环、Krebs循环),(乙酰CoA H2O 和CO2,释放出能量)
,(乙酰CoA H2O 和CO2,释放出能量)")
25
(一)反应过程 1、丙酮酸氧化脱羧生成乙酰CoA:线粒体 丙酮酸脱氢酶复合体(线粒体内膜) p345 丙酮酸脱氢酶(E1) 、二氢硫辛酰转乙酰基酶 (E2)、二氢硫辛酸脱氢酶(E3)。 辅助因子:HSCoA,焦磷酸硫胺素(TPP),硫辛酸, NAD+, FAD,Mg2+。
 、二氢硫辛酰转乙酰基酶 (E2)、二氢硫辛酸脱氢酶(E3)。 辅助因子:HSCoA,焦磷酸硫胺素(TPP),硫辛酸, NAD+, FAD,Mg2+。 .")
26
P348图10-7

27
2、乙酰CoA进入三羧酸循环彻底氧化 ① 三羧酸循环的反应过程: 9步(P350图10-8) Ⅰ. 缩合反应 * 柠檬酸合酶 ▲
 Ⅰ. 缩合反应 * 柠檬酸合酶 ▲")
28
Ⅱ.第一次氧化脱羧 * ▲ * ▲

29
Ⅲ.第二次氧化脱羧 α-酮戊二酸脱氢酶复合体: α-酮戊二酸脱氢酶(E1) 、二氢硫辛酰转琥珀酰酶 (E2)、二氢硫辛酸脱氢酶(E3)。
* ▲ α-酮戊二酸脱氢酶复合体: α-酮戊二酸脱氢酶(E1) 、二氢硫辛酰转琥珀酰酶 (E2)、二氢硫辛酸脱氢酶(E3)。 辅助因子:HSCoA,焦磷酸硫胺素(TPP),硫辛酸, NAD+, FAD,Mg2+。
 、二氢硫辛酰转琥珀酰酶 (E2)、二氢硫辛酸脱氢酶(E3)。 辅助因子:HSCoA,焦磷酸硫胺素(TPP),硫辛酸, NAD+, FAD,Mg2+。")
30
概 念:在代谢过程中由于底物分子内部能量重新分布产生高能键而使ADP(GDP)磷酸化为ATP(GTP)的反应。
Ⅳ.底物水平磷酸化 * 琥珀酰CoA合成酶 概 念:在代谢过程中由于底物分子内部能量重新分布产生高能键而使ADP(GDP)磷酸化为ATP(GTP)的反应。 高能键的形成:脱氢、脱水、氧化脱羧。
磷酸化为ATP(GTP)的反应。 高能键的形成:脱氢、脱水、氧化脱羧。")
31
Ⅴ.琥珀酸氧化成草酰乙酸 * TCA的总反应:

32
TCA的总过程:

33
②.TCA的特点 Ⅰ. 四次脱氢:异柠檬酸氧化脱羧 -酮戊二酸氧化脱羧 3NADH+H+ 苹果酸氧化
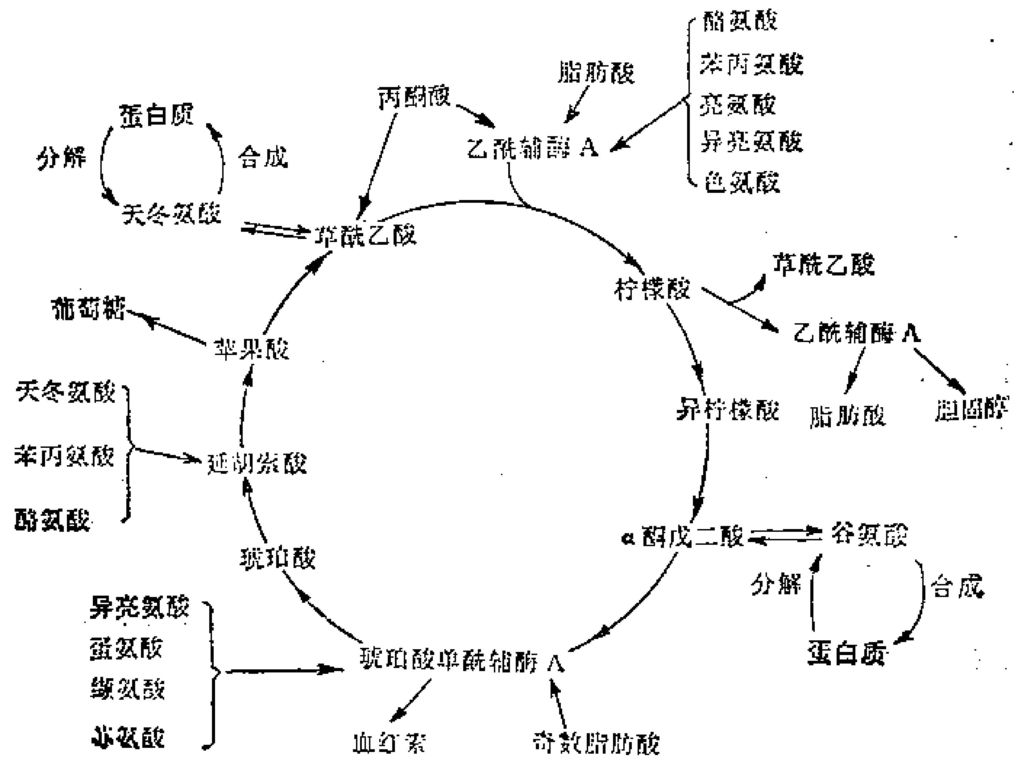
琥珀酸氧化 FADH2 两次脱羧:2CO2 Ⅱ. 有氧条件下进行,是产生ATP的主要途径 (p107) 3NADH+H FADH 底物水平 3 X 2.5ATP ATP GTP=10ATP Ⅲ.细胞部位:线粒体。 Ⅳ. TCA是不能逆行的 限速酶:柠檬酸合酶,异柠檬酸脱氢酶, -酮戊二酸脱氢酶复合体. Ⅴ. TCA中间产物的补充是必需的。 Ⅵ. TCA的生理意义 是三大营养物质分解代谢的最后共同途径,代谢枢纽,是获得能量最多的阶段,其中间产物是生物合成的前体来源。(p353)
 3NADH+H+ 1FADH2 底物水平 3 X 2.5ATP + 1.5ATP + 1GTP=10ATP Ⅲ.细胞部位:线粒体。 Ⅳ. TCA是不能逆行的 限速酶:柠檬酸合酶,异柠檬酸脱氢酶, -酮戊二酸脱氢酶复合体. Ⅴ. TCA中间产物的补充是必需的。 Ⅵ. TCA的生理意义. 是三大营养物质分解代谢的最后共同途径,代谢枢纽,是获得能量最多的阶段,其中间产物是生物合成的前体来源。(p353)")
34
P344,353 1 2.5×2* 1×2 2.5×2 1×2 1.5×2 2.5×2 28(26) 2 6
 2 6")
35
草酰乙酸的补充反应(回补反应) * 丙酮酸羧化(动物、酵母) * 磷酸烯醇式丙酮酸→草酰乙酸(植物、微生物) +CO2 +H2O
羧化酶 + H3PO4

36
丙酮酸 + CO2+ NADPH + H+ → 苹果酸 + NADP+
* 苹果酸酶的作用(动物、植物、微生物) 丙酮酸 + CO2+ NADPH + H+ → 苹果酸 + NADP+ 苹果酸酶 苹果酸+ NAD+ → 草酰乙酸+ NADH + H+ 苹果酸脱氢酶 *天门冬氨酸→草酰乙酸 *乙醛酸循环途径(植物和微生物) 2乙酰CoA+2H2O+ NAD+ → 琥珀酸+ 2CoASH+NADH + H+ 琥珀酸→→→草酰乙酸
 丙酮酸 + CO2+ NADPH + H+ → 苹果酸 + NADP+ 苹果酸酶. 苹果酸+ NAD+ → 草酰乙酸+ NADH + H+ 苹果酸脱氢酶. *天门冬氨酸→草酰乙酸. *乙醛酸循环途径(植物和微生物) 2乙酰CoA+2H2O+ NAD+ → 琥珀酸+ 2CoASH+NADH + H+ 琥珀酸→→→草酰乙酸.")
38
③ TCA的调节 NADH/NAD+ ATP/ADP 柠檬酸合酶 异柠檬酸脱氢酶 -酮戊二酸脱氢酶 TCA [草酰乙酸]在细胞较低,其浓度变化对TCA速度影响较大。 [Ca2+ ] 线 两种酶 TCA

40
三、糖的磷酸戊糖途径 1、概念: 是一条需氧的磷酸己糖氧化途径,又称为磷酸己糖支路(HMS)。以6-P-G为起始物,通过转醛基,转酮基等反应生成了 3C~7C等糖衍生物,是体内产生核糖(5C)和NADPH+H+的代谢途径。 2、细胞部位:胞液 3、反应过程: (1)氧化阶段(脱氢、脱羧) 6-P-G 5-磷酸核酮糖,生成 2NADPH、1CO2 (2)非氧化阶段(转酮、转醛反应) 一系列的基团转移反应,戊糖分子发生结合与重排, 再生出磷酸葡萄糖。
氧化阶段(脱氢、脱羧) 6-P-G 5-磷酸核酮糖,生成 2NADPH、1CO2 (2)非氧化阶段(转酮、转醛反应) 一系列的基团转移反应,戊糖分子发生结合与重排, 再生出磷酸葡萄糖。")
42
5-P-核酮糖 5-P-木酮糖 5-P-核糖 5-P-木酮糖 5-P-木酮糖 5-P-核糖 5-P-木酮糖 C3 C7 C4 C3 C3

43
×6 6 6NADPH+6H+ 6NADP+ 2× ×2 6-磷酸果糖 5×6-磷酸葡萄糖 脱氢酶 内脂酶 异构酶 差向酶 转酮酶 转醛酶

44
4、特点 (1)起始物:6-P- G 产物:3C~7C, NADPH+H+ (2)转酮基、转醛基反应 (3)关键酶:6-磷酸葡萄糖脱氢酶, 别构制:NADPH (4)反应式: G-6-P + 12NADP++7H20→6CO2+12NADPH+12H++H3PO4 5、意义 (1)是不同结构糖分子的重要来源。 (2)核糖是合成核酸、游离核苷酸、辅酶的原料 (3)NADPH+H+参与许多代谢反应 ◆ 合成代谢的供氢体:脂肪酸,胆固醇的合成 ◆ 参与羟化反应( O2、NADPH ):生物合成、生物转化 ◆ NADPH维持GSH于还原态,保护一些含-SH的蛋白质(酶)免受氧化剂,尤其是过氧化物(H2 O2)的损害。 NO 053
是不同结构糖分子的重要来源。 (2)核糖是合成核酸、游离核苷酸、辅酶的原料 (3)NADPH+H+参与许多代谢反应 ◆ 合成代谢的供氢体:脂肪酸,胆固醇的合成 ◆ 参与羟化反应( O2、NADPH ):生物合成、生物转化. ◆ NADPH维持GSH于还原态,保护一些含-SH的蛋白质(酶)免受氧化剂,尤其是过氧化物(H2 O2)的损害。 NO 053.")
45
四、其他分解代谢途径 1、糖醛酸途径(glucuromic acid cycle)
从G-6-P或G-1-P开始经UDP-葡萄糖生成葡萄糖醛酸的途径,进而可以合成抗坏血酸和5-磷酸木酮糖,后者与磷酸戊糖途径相联系。 2、乙醛酸循环(glyoxylate cycle) (1)微生物、植物所特有的TCA cycle 的变化形式, 是一个有机酸代谢环。 乙酰CoA→→异柠檬酸(异柠檬酸裂合酶)→乙醛酸+琥珀酸 +乙酰CoA →苹果酸(苹果酸合酶)→草酰乙酸(→磷酸烯醇式丙酮酸 → → →葡萄糖) 反应式: 2乙酰CoA+2NAD++FAD→草酰乙酸+2CoASH+2NADH+2H++FADH2
 (1)微生物、植物所特有的TCA cycle 的变化形式, 是一个有机酸代谢环。 乙酰CoA→→异柠檬酸(异柠檬酸裂合酶)→乙醛酸+琥珀酸. +乙酰CoA. →苹果酸(苹果酸合酶)→草酰乙酸(→磷酸烯醇式丙酮酸. → → →葡萄糖) 反应式: 2乙酰CoA+2NAD++FAD→草酰乙酸+2CoASH+2NADH+2H++FADH2.")
46
(2)是油料植物将脂肪酸转变为糖的途径(种子发芽时该途径很活跃)。
(3)是微生物利用乙酸作为碳源合成机体的途径。
是微生物利用乙酸作为碳源合成机体的途径。")
47
第三节 糖 异 生 一、概 述 1. 糖异生(Gluconeogenesis)的定义 :指生物体利用非糖物质转变成糖或糖原的过程。
第三节 糖 异 生 一、概 述 1. 糖异生(Gluconeogenesis)的定义 :指生物体利用非糖物质转变成糖或糖原的过程。 2.原料:甘油、有机酸(丙酮酸、乳酸、丙酸、TCA中的羧酸)、 生糖氨基酸。 3.部位:肝、肠、肾(胞液+线粒体) 4 . 酶:葡萄糖-6-磷酸酶、果糖-1,6-二磷酸酶、 丙酮酸羧化酶、磷酸烯醇式丙酮酸羧激酶
的定义 :指生物体利用非糖物质转变成糖或糖原的过程。 2.原料:甘油、有机酸(丙酮酸、乳酸、丙酸、TCA中的羧酸)、 生糖氨基酸。 3.部位:肝、肠、肾(胞液+线粒体) 4 . 酶:葡萄糖-6-磷酸酶、果糖-1,6-二磷酸酶、 丙酮酸羧化酶、磷酸烯醇式丙酮酸羧激酶.")
48
生糖氨基酸: Ala, Cys, Gly, Ser, Thr, Trp→ 丙酮酸 Pro,His,Gln,Arg→ Glu→ α-酮戊二酸 Ile,Met,Ser,Thr,Val→ 琥珀酰CoA Phe,Tyr→ 延胡索酸 Asn,Asp→ 草酰乙酸 甘油: 甘油三酯→甘油→α-磷酸甘油→磷酸二羟丙酮。 乳酸: 乳酸→丙酮酸。

49
例:甘油、乳酸异生成糖 p373图10-22

50
1.丙酮酸 磷酸烯醇型丙酮酸(丙酮酸羧化支路)
二、反应过程(与酵解比较) 1.丙酮酸 磷酸烯醇型丙酮酸(丙酮酸羧化支路) ● 线粒体
 1.丙酮酸 磷酸烯醇型丙酮酸(丙酮酸羧化支路) ● 线粒体.")
51
2. 1,6一二磷酸果糖 6一磷酸果糖

52
葡萄糖-6-磷酸酶不存在于脑和肌肉组织中,故脑和肌肉组织不能生成自由葡萄糖。
3.6-磷酸葡萄糖水解为葡萄糖 葡萄糖-6-磷酸酶不存在于脑和肌肉组织中,故脑和肌肉组织不能生成自由葡萄糖。

53
三、生 理 意 义 1、空腹、饥饿、剧烈运动,维持血糖水平恒定 G→
脂肪分解甘油 G 饥饿 Pr.分解生糖AA → G 脑、红细胞 G 2、异生能量消耗 丙酮酸羧化 ATP 草酰乙酸→ 磷酸烯醇式丙酮酸 GTP 3×2=6 3-磷酸甘油酸→ 1,3-二磷酸甘油酸 ATP 2分子丙酮酸→1分子G,消耗6分子ATP。 1、空腹、饥饿、剧烈运动,维持血糖水平恒定 G→

54
3、回收乳酸分子中的能量:乳酸循环 (Cori cycle)
乳酸的再利用,肝糖原更新,补充肌肉消耗的糖及防止乳酸酸中毒。 4、维持酸碱平衡

55
三、糖异生的调节(与糖酵解的协调) 1、代谢物
丙酮酸羧化酶 丙酮酸激酶 果糖-1,6-二磷酸酶 磷酸果糖激酶-1 糖异生 ATP、柠檬酸 ADP、AMP 果糖-2,6-二磷酸 另: 丙酮酸脱氢酶 丙酮酸羧化酶 乙酰CoA

56
2、激素 升糖激素:肾上腺糖皮质激素、肾上腺素、胰高血糖素等 作用:抑制酵解、增加糖异生。 机制:1)使糖异生途径的某些酶活性增高或合成量增多,如磷酸烯醇式丙酮酸羧激酶,葡萄糖-6-磷酸酶合成增加;2)通过磷酸化作用,改变酶活性,如磷酸果糖激酶、丙酮酸激酶磷酸化后活力降低,酵解受抑制,果糖二磷酸酶磷酸化后活力升高。 降糖激素:胰岛素 作用:增加酵解,抑制糖异生,使血糖降低。 机制:改变酶活性 ,使肝磷酸果糖激酶、丙酮酸激酶活性升高,酵解增强,果糖二磷酸酶活力下降,糖异生减弱。
使糖异生途径的某些酶活性增高或合成量增多,如磷酸烯醇式丙酮酸羧激酶,葡萄糖-6-磷酸酶合成增加;2)通过磷酸化作用,改变酶活性,如磷酸果糖激酶、丙酮酸激酶磷酸化后活力降低,酵解受抑制,果糖二磷酸酶磷酸化后活力升高。 降糖激素:胰岛素 作用:增加酵解,抑制糖异生,使血糖降低。 机制:改变酶活性 ,使肝磷酸果糖激酶、丙酮酸激酶活性升高,酵解增强,果糖二磷酸酶活力下降,糖异生减弱。")
57
第四节 糖原的合成与分解 糖原(glycogen): 动物细胞葡萄糖的储存形式 肌糖原:180-300g 肌肉收缩的应急能源 种类
第四节 糖原的合成与分解 糖原(glycogen): 动物细胞葡萄糖的储存形式 肌糖原: g 肌肉收缩的应急能源 种类 肝糖原:70-100g 血糖
: 动物细胞葡萄糖的储存形式. 肌糖原: g 肌肉收缩的应急能源. 种类. 肝糖原:70-100g 血糖.")
58
糖原是一种无还原性的多糖。 糖原合成或分解时,其葡萄糖残基的添加或去除,均在其非还原端进行。 糖原的合成与分解代谢主要发生在肝、肾和肌肉组织细胞的胞液中。 糖原合成与分解的生理意义: (1)贮存和动员能量。 (2)调节血糖浓度。 (3)利用乳酸:肝中可经糖异生途径利用糖无氧酵解产生的乳酸来合成糖原。这就是肝糖原合成的三碳途径或间接途径。
调节血糖浓度。 (3)利用乳酸:肝中可经糖异生途径利用糖无氧酵解产生的乳酸来合成糖原。这就是肝糖原合成的三碳途径或间接途径。")
59
一、糖原的分解代谢

60
* 1、反应过程: 分为三个阶段 (1)水解:包括三步反应,循环交替进行。
1)磷酸解:由糖原磷酸化酶催化对α-1,4-糖苷键磷酸解,生成G-1-P。 * 糖原磷酸化酶 (G)n + Pi (G)n-1 + G-1-P
磷酸解:由糖原磷酸化酶催化对α-1,4-糖苷键磷酸解,生成G-1-P。 * 糖原磷酸化酶. (G)n + Pi (G)n-1 + G-1-P.")
61
由葡萄糖-6-磷酸酶催化,生成自由葡萄糖,该酶只存在于肝及肾中。
2)异构: 磷酸葡萄糖变位酶 G-1-P G-6-P 3)脱磷酸: 由葡萄糖-6-磷酸酶催化,生成自由葡萄糖,该酶只存在于肝及肾中。 葡萄糖-6-磷酸酶 G-6-P + H2O G + Pi
异构: 磷酸葡萄糖变位酶. G-1-P G-6-P. 3)脱磷酸: 由葡萄糖-6-磷酸酶催化,生成自由葡萄糖,该酶只存在于肝及肾中。 葡萄糖-6-磷酸酶. G-6-P + H2O G + Pi.")
62
(2)转寡糖链:当糖原被水解到离分支点四个葡萄糖残基时,由葡聚糖转移酶催化,将分支链上的三个葡萄糖残基转移到直链的非还原端,使分支点暴露。
(3)脱支:由α-1,6-葡萄糖苷酶催化。将α-1,6-糖苷键水解,生成一分子自由葡萄糖。 α-1,6-葡萄糖苷酶 (G)n + H2O (G)n-1 + G
脱支:由α-1,6-葡萄糖苷酶催化。将α-1,6-糖苷键水解,生成一分子自由葡萄糖。 α-1,6-葡萄糖苷酶. (G)n + H2O (G)n-1 + G.")
63
2、糖原分解的特点: 1.水解反应在糖原的非还原端进行; 2.是一非耗能过程;
3.关键酶是糖原磷酸化酶,为一共价修饰酶,其辅酶是磷酸吡哆醛。

64
二、糖原的合成代谢 1、反应过程: 分为三个阶段 (1)活化:由葡萄糖生成UDPG,是一耗能过程。 1) 磷酸化:
己糖激酶(葡萄糖激酶) G + ATP G-6-P + ADP 2) 异构: 磷酸葡萄糖变位酶 G-6-P G-1-P 3) 转形:G-1-P转变为尿苷二磷酸葡萄糖(UDPG): UDPG焦磷酸化酶 G-1-P + UTP UDPG + PPi
 G + ATP G-6-P + ADP. 2) 异构: 磷酸葡萄糖变位酶. G-6-P G-1-P. 3) 转形:G-1-P转变为尿苷二磷酸葡萄糖(UDPG): UDPG焦磷酸化酶. G-1-P + UTP UDPG + PPi.")
65
* (2)缩合: UDPG + (G)n (G)n+1 + UDP (3)分支:
糖原合酶 * (3)分支: 当直链长度达12个葡萄糖残基以上时,在分支酶的催化下,将距末端6~7个葡萄糖残基组成的寡糖链由α-1,4-糖苷键转变为α-1,6-糖苷键,使糖原出现分支。
分支: 当直链长度达12个葡萄糖残基以上时,在分支酶的催化下,将距末端6~7个葡萄糖残基组成的寡糖链由α-1,4-糖苷键转变为α-1,6-糖苷键,使糖原出现分支。")
66
α-1,4 α-1,6

67
2、糖原合成的特点: 1.必须以原有糖原分子作为引物; 2.合成反应在糖原的非还原端进行;
3.合成为一耗能过程,每增加一个葡萄糖残基,需消耗2个高能磷酸键(2分子ATP); 4.其关键酶是糖原合酶,为一共价修饰酶; 5.需UTP参与(以UDP为载体)。
; 4.其关键酶是糖原合酶,为一共价修饰酶; 5.需UTP参与(以UDP为载体)。")
68
三、糖原合成与分解的调节 糖原合成:糖原合酶 糖原分解:磷酸化酶 1.共价修饰(磷酸化和去磷酸化) 磷酸化 去磷酸化
糖原合成:糖原合酶 糖原分解:磷酸化酶 1.共价修饰(磷酸化和去磷酸化) 磷酸化 去磷酸化 磷酸化酶a(活性形式) 磷酸化酶b(无活性形式) 糖原合酶b(无活性形式) 糖原合酶a(活性形式) 糖原降解 糖原合成 2.别构调节 别构抑制剂 糖原的分解 磷酸化酶a ATP ,6-P-G 别构激活剂 磷酸化酶b AMP 别构激活剂 糖原的合成 糖原合酶b P-G
 磷酸化 去磷酸化. 磷酸化酶a(活性形式) 磷酸化酶b(无活性形式) 糖原合酶b(无活性形式) 糖原合酶a(活性形式) 糖原降解 糖原合成. 2.别构调节 别构抑制剂 糖原的分解 磷酸化酶a ATP ,6-P-G 别构激活剂 磷酸化酶b AMP 别构激活剂 糖原的合成 糖原合酶b 6-P-G ")
69
糖原合酶b 糖原合酶a

70
第五节 光合作用 光合作用细胞 异养细胞 糖 太阳能是所有生物能的最终来源

71
光合作用(Photosynthesis)是糖合成代谢的主要途径。据估计,每年地球上约有1011吨CO2的碳被固定。
绿色植物、光合细菌或藻类等将光能转变成化学能的过程,即利用光能,由CO2和H2O合成糖类化合物并释放出氧气的过程,称为光合作用。 光合作用使得地球上的碳构成一个碳循环,即非光养生物通过氧化糖获得能量,并释放出CO2,而光养生物捕获CO2并使其还原为糖。 光合作用的总反应式可表示如下: 光能 nCO nH2O (CH2O)n + nO2 + nH2O 叶绿体
n + nO2 + nH2O. 叶绿体.")
72
一、叶绿体及光合色素 1.叶绿体 植物的绿色部分含有叶绿体,是绿色植物进行光合作用的场所。 叶绿体由外膜和内膜组成,内外膜之间有膜间隙。膜内为基质,包含有许多可溶性酶,是进行暗反应的场所,是将二氧化碳还原为糖的部位。基质内分布着片层状类囊体,是一个高度折叠的、连续膜网络结构,含有大量可进行光反应的光合色素,是导致NADPH和ATP形成的光反应部位。

73
膜间空隙 外膜 内膜 基质 腔 基粒 基质片层 类囊体膜 基粒片层

74
2.叶绿素(chlorophyll,Chl)
绿色植物叶绿体中接受光能的主要组分是叶绿素,包括叶绿素a和叶绿素b,其它的光合色素是类胡萝卜素(400~500nm)等。光合细菌和藻类中还含有叶绿素c和藻胆色素(520~630nm)等。叶绿素分子通过共价和非共价键与内在膜蛋白结合,按特定的取向排列于光合作用膜上。 叶绿素是一类含镁的卟啉衍生物,带羧基的侧链与一个含有20个碳的叶绿醇形成酯,该侧链有助于叶绿素融解在膜中。 叶绿素不溶于水,能溶于有机溶剂,在可见光区有很强的吸收,叶绿素a为680 nm, 叶绿素b为460 nm。
等。光合细菌和藻类中还含有叶绿素c和藻胆色素(520~630nm)等。叶绿素分子通过共价和非共价键与内在膜蛋白结合,按特定的取向排列于光合作用膜上。 叶绿素是一类含镁的卟啉衍生物,带羧基的侧链与一个含有20个碳的叶绿醇形成酯,该侧链有助于叶绿素融解在膜中。 叶绿素不溶于水,能溶于有机溶剂,在可见光区有很强的吸收,叶绿素a为680 nm, 叶绿素b为460 nm。")
76
二、光反应 (1)光反应系统 光反应过程是由叶绿素的两种光合系统(photosystem),即光系统I (PS I) 和光系统II (PS II) 共同完成的,又被称为光反应中心,存在于叶绿体的类囊体膜中。
,即光系统I (PS I) 和光系统II (PS II) 共同完成的,又被称为光反应中心,存在于叶绿体的类囊体膜中。 .")
77
光系统II (PS II) PSII主要位于基粒片层中,远离基质,PS II有一个能够捕获光能的复合体, 是由大约200个叶绿素分子、50个类胡萝卜素分子以及12条多肽链等组成的跨膜复合物。光能首先被该系统的色素分子所吸收,所以常称为天线色素。 反应中心含有50个叶绿素a,以及质体醌等电子供体和受体。由天线色素吸收的光能以激发能形式转移入反应中心,并产生一种强氧化剂和一种弱还原剂。由于反应中心在波长680 nm 处有最大吸收,又称为P680 (P指色素,680是最大吸收波长nm)。 放氧复合体含有能促进水裂解的蛋白(含有Mn2+离子)等。反应中心产生的强氧化剂在水裂解酶催化下,将水裂解成氧和电子,这高能电子是推动暗反应的动力。
。 放氧复合体含有能促进水裂解的蛋白(含有Mn2+离子)等。反应中心产生的强氧化剂在水裂解酶催化下,将水裂解成氧和电子,这高能电子是推动暗反应的动力。")
78
光系统I (PS I) PS I是主要位于基质片层,暴露于叶绿体基质,是一个跨膜复合物,由13条多肽链及200个叶绿素、50个类胡萝卜素以及细胞色素bf、质体蓝素(简写为PC)和铁氧还蛋白(简写为FD)等组成。PS I 的反应中心含有130个叶绿素a,它的最大吸收波长为700 nm,所以又称为P700。 PS I 在波长为700 nm的光照下被激活,产生一种强还原剂和一种弱氧化剂。强还原剂在铁氧还蛋白作用下,生成NADPH,是暗反应的主要还原剂。
和铁氧还蛋白(简写为FD)等组成。PS I 的反应中心含有130个叶绿素a,它的最大吸收波长为700 nm,所以又称为P700。 PS I 在波长为700 nm的光照下被激活,产生一种强还原剂和一种弱氧化剂。强还原剂在铁氧还蛋白作用下,生成NADPH,是暗反应的主要还原剂。")
79
光合作用成分在类囊体膜中的分布

80
除了光系统和LHC(叶绿素a/b集光复合体)之外,类囊体膜上或是与它相连接的成分也参与光合作用,其中包括水裂解复合体(促氧生成复合体)、细胞色素bf复合体和叶绿体ATP合酶。
水裂解复合体是由几个外周蛋白质和四个锰原子组成的,与位于类囊体膜边上的PSII相联。锰原子直接与反应中心的蛋白质结合,构成了活性中心。外周蛋白与反应中心非共价结合,在生成氧的反应中起协助作用。细胞色素bf复合体跨越类囊体膜,位于基质和基粒片层中。ATP合酶也跨越类囊体膜,但只位于基质片层中。 在光能的驱动下类囊体内进行着电子和质子流动。在水裂解复合体的作用下,来自H2O裂解产生的电子经PSII、一系列中间载体、PSI,最后传递给NADP+。质子在电子传递过程中从基质泵入类囊体腔内,与H2O裂解生成的质子在腔内外形成一个质子梯度,腔内的质子经ATP合酶形成的质子通道重新再流回基质,同时催化ATP的合成。

81
类囊体膜的光合作用成分及电子和质子的流动

82
(2)光反应的电子传递链(光合链) 光反应中心的色素分子P吸收一个光子,即形成激发态P*。激发态P*的电子具有很高的能量,是良好的电子供体,因此P*是一个强还原剂(PSI) 。 反应中心的叶绿素容易将一个电子传递给脱镁叶绿素,反应中心转变为P+的氧化作用和脱镁叶绿素转变为Ph-的还原作用而发生的电荷转移,是光能转换为化学能的重要步骤。失去了电子的P+,是一个好的电子受体,是一个强氧化剂(PSII)。 从P*释放出来的高能电子将沿着类囊体膜中的电子传递链传递,涉及两个光反应系统。光合链反应过程可分为两个阶段。
。 从P*释放出来的高能电子将沿着类囊体膜中的电子传递链传递,涉及两个光反应系统。光合链反应过程可分为两个阶段。")
83
电子传递链

84
(3) 光合磷酸化 通过光激发导致电子传递与磷酸化作用相偶联合成ATP的过程,称为光合磷酸化。按照光合链电子传递的方式,光合磷酸化可以分为两种形式。 非环式光合磷酸化:在光照条件下,水分子光裂解产生的电子,经P680将电子传递到NADP+,电子流动经过两个光系统,两次被激发成高能电子。电子传递过程中产生的质子梯度,驱动ATP合成,并生成NADPH。 环式光合磷酸化:PS I作用中心P700受光激发释放出的高能电子, 在传递到铁氧还蛋白后,不再继续向NADP+传递,而是将电子传回给细胞色素bf复合物,然后细胞色素bf又将电子通过质体蓝素传递给P700。电子在此循环流动过程中,产生质子梯度,从而驱动ATP的合成。环式光合磷酸化只涉及PS I,并且只生成ATP而无NADPH生成、无O2释放。

85
三、暗反应 暗反应是指由光反应产生的NADPH在ATP供给能量情况下,将CO2还原成糖的反应过程。这是一个酶催化的反应过程,不需要光参加。
大多数植物的暗反应中,还原CO2的第一个产物是三碳化合物(3-磷酸甘油酸),所以这种途径称为C3途径。有些植物,如甘蔗和玉米等高产作物,其暗反应还原CO2的产物是四碳化合物(草酰乙酸等),所以称为C4途径。
,所以这种途径称为C3途径。有些植物,如甘蔗和玉米等高产作物,其暗反应还原CO2的产物是四碳化合物(草酰乙酸等),所以称为C4途径。")
86
C3途径 C3途径的反应以循环形式进行,又称为三碳循环,也称为Calvin循环,还原戊糖磷酸(reductive pentose phosphate,RPP)循环。 这个阶段包括三个主要的过程:一是CO2受体固定大气中CO2;二是将固定的CO2还原为糖;三是可接受CO2的受体分子的重新生成。
循环。 这个阶段包括三个主要的过程:一是CO2受体固定大气中CO2;二是将固定的CO2还原为糖;三是可接受CO2的受体分子的重新生成。")
88
能量消耗 上述所有反应组成了一个循环.每一个循环,1分子的二磷酸核酮糖固定1分子CO2,生成1/6分子6-磷酸果糖,其中5/6分子的6-磷酸果糖参与再循环,1/6分子的6-磷酸果糖则转变成葡萄糖。从CO2的固定到生成一分子葡萄糖共需6个循环,总反应式是: 6 CO H ATP + 12 NADPH + 12 H2O C6H12O ADP + 12 NADP+ + 6 H+ G’ = kJ / mol 上式表明,在三碳循环中,每还原1分子CO2需要消耗3分子ATP和2分子NADPH。

89
C4途径 C4途径分为羧化和脱羧两个阶段。在起始阶段,CO2在叶肉细胞被固定形成C4酸。第二阶段C4酸在维管束鞘细胞中脱羧,释放出的CO2再被固定,进入RPP循环。

91
本 章 小 结 糖的转变概况:糖的消化(降解)、吸收、转运及中间代谢过程,包括分解代谢和合成代谢。血糖的来源与去路。
糖的代谢途径:生物机体中重要的代谢途径,包括糖酵解(发酵)、糖的有氧氧化(三羧酸循环)、磷酸戊糖途径、乙醛酸循环、糖异生途径、糖原的分解与合成、光合作用等。相关途径的组织、细胞定位,反应过程(关键反应、限速反应),关键酶(限速酶),特点,生理意义,主要的调节物和调节方式。能量产生和利用的部位。 名词解释:糖酵解(途径)、酵解与发酵、糖的有氧氧化、柠檬酸循环、底物水平磷酸化、乳酸循环、磷酸戊糖途径、糖异生、丙酮酸脱氢酶复合体、α-酮戊二酸脱氢酶复合体、光合作用、光合磷酸化、光反应、暗反应
、糖的有氧氧化(三羧酸循环)、磷酸戊糖途径、乙醛酸循环、糖异生途径、糖原的分解与合成、光合作用等。相关途径的组织、细胞定位,反应过程(关键反应、限速反应),关键酶(限速酶),特点,生理意义,主要的调节物和调节方式。能量产生和利用的部位。 名词解释:糖酵解(途径)、酵解与发酵、糖的有氧氧化、柠檬酸循环、底物水平磷酸化、乳酸循环、磷酸戊糖途径、糖异生、丙酮酸脱氢酶复合体、α-酮戊二酸脱氢酶复合体、光合作用、光合磷酸化、光反应、暗反应.")
92
思 考 题 1、比较底物水平磷酸化、光合磷酸化与氧化磷酸化三者的异同。 2、请指出血糖的来源与去路。
3、为什么说6-磷酸葡萄糖是各个糖代谢途径的交叉点。 4、结合激素的作用机制,说明肾上腺素如何通过对有关酶类的活性调节实现对血糖浓度的调控。

93
本章结束

Similar presentations






 反应部位 器官,细胞内定位 生理意义 代谢调节 主要调节点,主要变构抑制剂、变构激活剂 各代谢途径之间的联系和调控.>")


 合成代谢------需要能量 物质代谢 能量代谢>")



蛋白质的降解: 外源蛋白的消化 内源性蛋白的选择性降解 (2)氨基酸的分解代谢:>")





.>")





