Download presentation
Presentation is loading. Please wait.

1
第五章 糖及糖的分解代谢

2
新陈代谢是所有生物维持其生命活动的最基本的特性,是生物体内有机物合成和分解作用,包括物质转变和能量转化。
前言:新陈代谢的概念 新陈代谢是所有生物维持其生命活动的最基本的特性,是生物体内有机物合成和分解作用,包括物质转变和能量转化。 合成代谢 物质上---小分子---大分子 (同化作用)能量上---积能过程 生物体新陈代谢 分解代谢 物质上---大分子---小分子 (异化作用)能量上---放能过程
能量上---积能过程. 生物体新陈代谢. 分解代谢 物质上---大分子---小分子. (异化作用)能量上---放能过程.")
3
新陈代谢就是在合成和分解过程中不断就得平衡。若合成大于分解,生命体旺盛;反之,则衰老。
前言:新陈代谢的概念 新陈代谢就是在合成和分解过程中不断就得平衡。若合成大于分解,生命体旺盛;反之,则衰老。

4
新陈代谢就是与糖类的分解有密切的联系,因为糖类的分解对生物体来讲,具重要的意义。
前言:新陈代谢的概念 新陈代谢就是与糖类的分解有密切的联系,因为糖类的分解对生物体来讲,具重要的意义。 1. 糖类作为能源物质 生物细胞的各种代谢活动,包括物质分解和合成都需要有足够的能量,其中ATP是糖类降解时通过氧化磷酸化作用而形成的最重要的能量载体物质。生物细胞只能利用高能化合物(主要是ATP)水解时释放的化学能来做功,以满足生长发育等所需要的能量消耗。
水解时释放的化学能来做功,以满足生长发育等所需要的能量消耗。")
5
前言:新陈代谢的概念 2.作为合成生物体内重要代谢物质的碳架和前体 葡萄糖、果糖等在降解过程中除了能提供大量能量外,其分解过程中还能形成许多中间产物或前体,生物细胞通过这些前体产物再去合成一系列其它重要的物质,包括: (1) 乙酰辅酶A、氨基酸、核苷酸等,它们分别是合成脂肪、蛋白质和核酸等大分子物质的前体。 (2) 生物体内许多重要的次生代谢物、抗性物质,如生物碱、黄酮类等物质,它们对提高植物的抗逆性起着重要的作用。
 乙酰辅酶A、氨基酸、核苷酸等,它们分别是合成脂肪、蛋白质和核酸等大分子物质的前体。 (2) 生物体内许多重要的次生代谢物、抗性物质,如生物碱、黄酮类等物质,它们对提高植物的抗逆性起着重要的作用。")
6
前言:新陈代谢的概念 3. 细胞中结构物质 细胞中的结构物质如植物细胞壁等是由纤维素、半纤维素、果胶质等物质组成;甲壳质或几丁质为N-乙酰葡萄糖胺的同聚物,是组成虾、蟹、昆虫等外骨骼的结构物质。这些物质都是由糖类转化物聚合而成。

7
前言:新陈代谢的概念 4. 参与分子和细胞特异性识别 由寡糖或多糖组成的糖链常存在于细胞表面,形成糖脂和糖蛋白,参与分子或细胞间的特异性识别和结合,如抗体和抗原、激素和受体、病原体和宿主细胞、蛋白质和抑制剂等常通过糖链识别后再进行结合。

8
第一节 重要糖类结构和双糖、多糖的降解 单糖(monosaccharide)是指最简单的糖,即在温和条件下不能再分解成更小的单体糖,如葡萄糖、果糖等。按碳原子的数目单糖又可分为三碳(丙)糖、四碳(丁)糖、五碳(戊)糖、六碳(已)糖、七碳(庚)糖等。
是指最简单的糖,即在温和条件下不能再分解成更小的单体糖,如葡萄糖、果糖等。按碳原子的数目单糖又可分为三碳(丙)糖、四碳(丁)糖、五碳(戊)糖、六碳(已)糖、七碳(庚)糖等。")
9
一、 一些重要单糖的结构 三 糖 甘油醛 二羟丙酮

10
一、 一些重要单糖的结构 四 碳 糖 赤藓糖

11
一、 一些重要单糖的结构 五 碳 糖 核 糖 核酮糖 木 糖

12
一、 一些重要单糖的结构 葡萄糖(glucose) ——已醛糖 果糖(fructose) ——已酮糖
 ——已醛糖 果糖(fructose) ——已酮糖")
13
一、 一些重要单糖的结构 D-7-磷酸-景天庚酮糖

14
二、 一些重要双糖的结构 α-葡萄糖(1→4)葡萄糖苷 麦芽糖
葡萄糖苷 麦芽糖")
15
二、 一些重要双糖的结构 α-葡萄糖(1→2)β-果糖苷 蔗 糖
β-果糖苷 蔗 糖")
16
二、 一些重要双糖的结构 乳糖(半乳糖--1, 4-葡萄糖) 乳 糖
 乳 糖")
17
三、 一些重要多糖的结构 直链: a-1,4-糖苷键 分支点: a-1,6-糖苷键 淀 粉

18
三、 一些重要多糖的结构 直链: a-1,4-糖苷键 分支点: a-1,6-糖苷键 淀 粉

19
三、 一些重要多糖的结构 b-1,4-糖苷键 纤 维 素

20
三、 一些重要多糖的结构 Chitin

21
四、淀粉的降解 淀粉分解有两条途径: 水解 → 产生葡萄糖 磷酸解 → 产生磷酸葡萄糖

22
参与淀粉水解的酶主要有三种:淀粉酶、脱支酶、 麦芽糖酶
1. 淀粉的水解 四、淀粉的降解 参与淀粉水解的酶主要有三种:淀粉酶、脱支酶、 麦芽糖酶 (1)淀粉酶: 淀粉酶是指参与淀粉a-1,4-糖苷键水解的酶。 有a-淀粉酶和b-淀粉酶两种。
淀粉酶: 淀粉酶是指参与淀粉a-1,4-糖苷键水解的酶。 有a-淀粉酶和b-淀粉酶两种。")
23
四、淀粉的降解 (1)淀粉酶: 1. 淀粉的水解 a-淀粉酶:(a-1,4-葡聚糖水解酶)
1. 淀粉的水解 四、淀粉的降解 (1)淀粉酶: a-淀粉酶:(a-1,4-葡聚糖水解酶) 可水解任何部位的a-1,4-糖苷键,所以又称为内切淀粉酶。 该酶对非还原末端的5个葡萄糖基不发生作用。Ca2+需要。 其产物为: 若直链淀粉 → 葡萄糖 + 麦芽糖 + 麦芽三糖 + 低聚糖 若支链淀粉 → 葡萄糖 + 麦芽糖 + 麦芽三糖 + 极限糊精
淀粉酶: a-淀粉酶:(a-1,4-葡聚糖水解酶) 可水解任何部位的a-1,4-糖苷键,所以又称为内切淀粉酶。 该酶对非还原末端的5个葡萄糖基不发生作用。Ca2+需要。 其产物为: 若直链淀粉 → 葡萄糖 + 麦芽糖 + 麦芽三糖 + 低聚糖. 若支链淀粉 → 葡萄糖 + 麦芽糖 + 麦芽三糖 + 极限糊精.")
24
四、淀粉的降解 (1)淀粉酶: β-淀粉酶: 其产物为: 若直链淀粉 → 麦芽糖 若支链淀粉 → 麦芽糖 + 极限糊精 1. 淀粉的水解
1. 淀粉的水解 四、淀粉的降解 (1)淀粉酶: β-淀粉酶: 也水解a-1,4-糖苷键,但须从非还原末端开始切,每次切下两个葡萄糖基。又称为外切淀粉酶。 其产物为: 若直链淀粉 → 麦芽糖 若支链淀粉 → 麦芽糖 + 极限糊精
淀粉酶: β-淀粉酶: 也水解a-1,4-糖苷键,但须从非还原末端开始切,每次切下两个葡萄糖基。又称为外切淀粉酶。 其产物为: 若直链淀粉 → 麦芽糖. 若支链淀粉 → 麦芽糖 + 极限糊精.")
25
(2)脱支酶(R-酶):(a-1,6-葡萄糖苷酶)
1. 淀粉的水解 四、淀粉的降解 (2)脱支酶(R-酶):(a-1,6-葡萄糖苷酶) 水解a-1,6-糖苷键,但只能作用于外围的这种键,而不能水解内部的分支。 (3)麦芽糖酶: 植物体内的麦芽糖酶通常与淀粉酶同时存在,并配合使用,从而使淀粉彻底水解成葡萄糖。
脱支酶(R-酶):(a-1,6-葡萄糖苷酶) 水解a-1,6-糖苷键,但只能作用于外围的这种键,而不能水解内部的分支。 (3)麦芽糖酶: 植物体内的麦芽糖酶通常与淀粉酶同时存在,并配合使用,从而使淀粉彻底水解成葡萄糖。")
26
Hydrolysis of glycogen and starch by -amylase and -amylase
1. 淀粉的水解 四、淀粉的降解 Hydrolysis of glycogen and starch by -amylase and -amylase

27
四、淀粉的降解 其中,淀粉磷酸化酶又叫P-酶。 (1)[Pi]很高(如施肥) (2)[G-1-P]低(因不断被利用)
2. 淀粉的磷酸解 四、淀粉的降解 其中,淀粉磷酸化酶又叫P-酶。 此反应为可逆反应,但在植物体内,由于 (1)[Pi]很高(如施肥) (2)[G-1-P]低(因不断被利用) 所以,反应向正方向进行。
![四、淀粉的降解 其中,淀粉磷酸化酶又叫P-酶。 (1)[Pi]很高(如施肥) (2)[G-1-P]低(因不断被利用)](http://slidesplayer.com/slide/11361085/61/images/27/%E5%9B%9B%E3%80%81%E6%B7%80%E7%B2%89%E7%9A%84%E9%99%8D%E8%A7%A3+%E5%85%B6%E4%B8%AD%EF%BC%8C%E6%B7%80%E7%B2%89%E7%A3%B7%E9%85%B8%E5%8C%96%E9%85%B6%E5%8F%88%E5%8F%ABP-%E9%85%B6%E3%80%82+%EF%BC%881%EF%BC%89%5BPi%5D%E5%BE%88%E9%AB%98%EF%BC%88%E5%A6%82%E6%96%BD%E8%82%A5%EF%BC%89+%EF%BC%882%EF%BC%89%5BG-1-P%5D%E4%BD%8E%EF%BC%88%E5%9B%A0%E4%B8%8D%E6%96%AD%E8%A2%AB%E5%88%A9%E7%94%A8%EF%BC%89.jpg "2. 淀粉的磷酸解. 四、淀粉的降解. 其中,淀粉磷酸化酶又叫P-酶。 此反应为可逆反应,但在植物体内,由于. (1)[Pi]很高(如施肥) (2)[G-1-P]低(因不断被利用) 所以,反应向正方向进行。")
28
2. 淀粉的磷酸解 四、淀粉的降解 淀粉磷酸化酶从淀粉的非还原端开始,一个一个地磷酸解a-1,4-糖苷键,直到距分支点4个葡萄糖基为止。 所以,如果是支链淀粉,还需要另外两个酶的参与,即转移酶和脱支酶。

29
2. 淀粉的磷酸解 四、淀粉的降解

30
2. 产物不易扩散到胞外,而水解产物葡萄糖会因扩散而流失
2. 淀粉的磷酸解 四、淀粉的降解 淀粉的磷酸解与水解相比,其优越性有: 1. 耗能少 2. 产物不易扩散到胞外,而水解产物葡萄糖会因扩散而流失

31
五、糖原的降解 糖原的磷酸解 糖原磷酸化酶(glycogen phosphorylase)是降解糖原的磷酸化的限速酶,有活性和非活性两种形式,分别称为糖原磷酸化酶a(活化态)和糖原磷酸化酶b(非活化态),两者在一定条件下可以相互转变。糖原磷酸解时,在磷酸化酶a作用下,从糖原非还原端开始逐个加磷酸切下葡萄糖生成1-磷酸葡萄糖,切至糖原分支点4个葡萄糖残基处为止。
是降解糖原的磷酸化的限速酶,有活性和非活性两种形式,分别称为糖原磷酸化酶a(活化态)和糖原磷酸化酶b(非活化态),两者在一定条件下可以相互转变。糖原磷酸解时,在磷酸化酶a作用下,从糖原非还原端开始逐个加磷酸切下葡萄糖生成1-磷酸葡萄糖,切至糖原分支点4个葡萄糖残基处为止。")
32
五、糖原的降解 糖原的磷酸解 转移酶(transferase)又称1, 41, 4葡聚糖转移酶,它主要作用是将连接与分支点上4个葡萄糖基的葡聚三糖转移至同一个分支点的另一个葡聚四糖链的末端,使分支点仅留下一个α(16)糖苷键连接的葡萄糖残基。
又称1, 41, 4葡聚糖转移酶,它主要作用是将连接与分支点上4个葡萄糖基的葡聚三糖转移至同一个分支点的另一个葡聚四糖链的末端,使分支点仅留下一个α(16)糖苷键连接的葡萄糖残基。 .")
33
五、糖原的降解 糖原的磷酸解 脱支酶,即水解α(16)糖苷键的酶,再将这个葡萄糖水解下来,使支链淀粉的分支结构变成直链结构,磷酸化酶再进一步将其降解为1-磷酸葡萄糖。由于磷酸化酶、转移酶和脱支酶的协同作用,将糖原(或支链淀粉)彻底降解。 糖原磷酸化酶主要存在于动物肝脏中,通过糖原分解直接补充血糖。
糖苷键的酶,再将这个葡萄糖水解下来,使支链淀粉的分支结构变成直链结构,磷酸化酶再进一步将其降解为1-磷酸葡萄糖。由于磷酸化酶、转移酶和脱支酶的协同作用,将糖原(或支链淀粉)彻底降解。 糖原磷酸化酶主要存在于动物肝脏中,通过糖原分解直接补充血糖。")
34
五、糖原的降解 The reactions of glycogen debranching enzyme

35
The structure of UDP-glucose, a sugar nucleotide
2. 形成糖核苷酸 六、蔗糖的降解 The structure of UDP-glucose, a sugar nucleotide

36
NDP主要是ADP和UDP,其产物分别为ADPG(腺苷二磷酸葡萄糖)和UDPG(尿苷二磷酸葡萄糖)。
2. 形成糖核苷酸 六、蔗糖的降解 NDP主要是ADP和UDP,其产物分别为ADPG(腺苷二磷酸葡萄糖)和UDPG(尿苷二磷酸葡萄糖)。 UDPG和ADPG是葡萄糖的活化形式,在合成寡糖和多糖时作为葡萄糖基的供体。这比将蔗糖水解要经济,因为从水解产物葡萄糖合成NDPG需要消耗能量。 蔗糖的这种降解方式在高等植物中普遍存在。 例如,在正在发育的谷类作物的籽粒能够将输入的蔗糖分解为ADPG,然后用以合成淀粉。
和UDPG(尿苷二磷酸葡萄糖)。 UDPG和ADPG是葡萄糖的活化形式,在合成寡糖和多糖时作为葡萄糖基的供体。这比将蔗糖水解要经济,因为从水解产物葡萄糖合成NDPG需要消耗能量。 蔗糖的这种降解方式在高等植物中普遍存在。 例如,在正在发育的谷类作物的籽粒能够将输入的蔗糖分解为ADPG,然后用以合成淀粉。")
37
第二节 糖酵解 glycolysis Louis Pasteur in his laboratory

38
(Embden-Meyerhof Parnas pathway)
一. 糖酵解的概念 定义 指葡萄糖通过一系列步骤,降解成三碳化合物(丙酮酸)的过程。 糖酵解途径又称 EMP途径 (Embden-Meyerhof Parnas pathway)
的过程。 糖酵解途径又称 EMP途径. (Embden-Meyerhof Parnas pathway)")
39
第一阶段 二. 糖酵解的过程 第一步:葡萄糖的磷酸化 激酶:催化将ATP上的磷酸基团转移到受体上的酶。激 酶都需要Mg2+作为辅助因子。

40
二. 糖酵解的过程

41
第一阶段 二. 糖酵解的过程 第二步:6-磷酸果糖的生成

42
二. 糖酵解的过程 第三步:1, 6-二磷酸果糖的生成 第一阶段
二. 糖酵解的过程 第一阶段 第三步:1, 6-二磷酸果糖的生成 磷酸果糖激酶(PFK)是EMP途径的关键酶,其活性大小控制着整个途径的进程。
是EMP途径的关键酶,其活性大小控制着整个途径的进程。")
43
Phosphofructokinase with ADP shown in white and fructose-6-P in red
二. 糖酵解的过程 Phosphofructokinase with ADP shown in white and fructose-6-P in red

44
碳链不变,但两头接上了磷酸基团,为断裂作好准备。
第一阶段 二. 糖酵解的过程 碳链不变,但两头接上了磷酸基团,为断裂作好准备。 消耗两个ATP。

45
第二阶段 二. 糖酵解的过程 第四步:1,6-二磷酸果糖的裂解 1个己糖分裂成2个丙糖 —— 丙酮糖和丙醛糖,它们为同分异构体。

46
第二阶段 二. 糖酵解的过程 第五步:磷酸丙糖的同分异构化 1分子二磷酸已糖裂解成2分子3-磷酸甘油醛。

47
二. 糖酵解的过程 第六步:3-磷酸甘油醛氧化 第三阶段
二. 糖酵解的过程 第六步:3-磷酸甘油醛氧化 糖酵解过程中第一次产生高能磷酸键,并且产生了还原剂 NADH。催化此反应的酶是巯基酶,所以它可被碘乙酸(ICH2COOH)不可逆地抑制。故碘乙酸能抑制糖酵解。
不可逆地抑制。故碘乙酸能抑制糖酵解。")
48
第三阶段 二. 糖酵解的过程 第七步:3-磷酸甘油酸和ATP的生成 糖酵解过程中第一次产生 ATP。

49
第三阶段 二. 糖酵解的过程 醛氧化成羧酸 NAD+还原成NADH 糖酵解中第一次产生ATP

50
第四阶段 二. 糖酵解的过程 第八步:3-磷酸甘油酸异构 Mg2+

51
第四阶段 二. 糖酵解的过程 The phosphoglycerate mutase of wheat germ catalyzes an intramolecular phosphoryl transfer

52
二. 糖酵解的过程 第九步:PEP的生成 Mg2+ 第四阶段
二. 糖酵解的过程 第九步:PEP的生成 Mg2+ 这一步其实是分子内的氧化还原,使分子中的能量重新分布,使能量集中,第二次产生了高能磷酸键。

53
第四阶段 二. 糖酵解的过程 第十步:丙酮酸的生成 Mg2+ 或 K+ 糖酵解过程中第二次产生 ATP。

56
葡萄糖 6-磷酸葡萄糖 6-磷酸果糖 1.6-二磷酸果糖 3磷酸甘油醛 磷酸二羟丙酮 1.3-二磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸
磷酸烯醇式丙酮酸 1.6-二磷酸果糖 丙酮酸 乳酸

57
三. 糖酵解的能量计算

58
要点: (1)反应部位:胞液 (2)关键酶:己糖激酶,6-磷酸果糖激酶-1,丙酮酸激酶 (3)能量的净生成:2ATP 消耗ATP的步骤: G
1,6-二磷酸果糖 生成ATP的步骤: 1,3-二磷酸甘油酸 ATP 3-磷酸甘油酸 磷酸烯醇式丙酮酸 丙酮酸

59
要点: (4)定位:细胞质 (5)意义:产生少许能量,产生一些中简产物如,丙酮酸和甘油等 (6)底物水平的磷酸化
定位:细胞质 (5)意义:产生少许能量,产生一些中简产物如,丙酮酸和甘油等 (6)底物水平的磷酸化")
60
糖酵解的调节 + 6-磷酸果糖激酶-1 6-磷酸果糖 ,6-二磷酸果糖 ADP ATP AMP 柠檬酸 2,6-二磷酸果糖 + -

61
6-磷酸果糖 2,6-二磷酸果糖 果糖二磷酸酶-2 6-磷酸果糖激酶-2 2,6-二磷酸果糖的合成与分解

62
磷酸烯醇式丙酮酸 丙酮酸激酶 长链脂肪酸 ATP 丙酮酸 1.6-二磷酸果糖 +

63
G 己糖激酶 6-磷酸葡萄糖

64
四. 糖酵解产物的去路 (1)在无氧或相对缺氧时 —— 发酵 有两种发酵:酒精发酵、乳酸发酵 酒精发酵:由葡萄糖 → 乙醇的过程
1. 丙酮酸的去路 四. 糖酵解产物的去路 (1)在无氧或相对缺氧时 —— 发酵 有两种发酵:酒精发酵、乳酸发酵 酒精发酵:由葡萄糖 → 乙醇的过程 丙酮酸脱羧酶需要TPP作为辅酶。
在无氧或相对缺氧时 —— 发酵. 有两种发酵:酒精发酵、乳酸发酵. 酒精发酵:由葡萄糖 → 乙醇的过程. 丙酮酸脱羧酶需要TPP作为辅酶。")
65
1. 丙酮酸的去路 四. 糖酵解产物的去路 (1)在无氧或相对缺氧时 ——酒精发酵
在无氧或相对缺氧时 ——酒精发酵")
66
四. 糖酵解产物的去路 (2)在无氧或相对缺氧时 ——乳酸发酵 乳酸发酵:由葡萄糖 → 乳酸的过程 乳酸脱氢酶在动物体内有5种同工酶:
1. 丙酮酸的去路 四. 糖酵解产物的去路 (2)在无氧或相对缺氧时 ——乳酸发酵 乳酸发酵:由葡萄糖 → 乳酸的过程 乳酸脱氢酶在动物体内有5种同工酶: H4、H3M、H2M2、HM3、M4
在无氧或相对缺氧时 ——乳酸发酵. 乳酸发酵:由葡萄糖 → 乳酸的过程. 乳酸脱氢酶在动物体内有5种同工酶: H4、H3M、H2M2、HM3、M4.")
67
许多微生物常进行这种过程。此外,高等动物在氧不充足时,也可进行这条途径,如肌肉强烈运动时即产生大量乳酸。
1. 丙酮酸的去路 四. 糖酵解产物的去路 (2)在无氧或相对缺氧时 ——乳酸发酵 许多微生物常进行这种过程。此外,高等动物在氧不充足时,也可进行这条途径,如肌肉强烈运动时即产生大量乳酸。
在无氧或相对缺氧时 ——乳酸发酵. 许多微生物常进行这种过程。此外,高等动物在氧不充足时,也可进行这条途径,如肌肉强烈运动时即产生大量乳酸。")
68
四. 糖酵解产物的去路 (3)在有氧条件下 —— 丙酮酸有氧氧化 丙酮酸被彻底氧化成CO2。 1. 丙酮酸的去路
四. 糖酵解产物的去路 (3)在有氧条件下 —— 丙酮酸有氧氧化 丙酮酸被彻底氧化成CO2。 这一过程在线粒体中进行。通过此过程可以使葡萄糖彻底降解、氧化成CO2。
在有氧条件下 —— 丙酮酸有氧氧化. 丙酮酸被彻底氧化成CO2。 这一过程在线粒体中进行。通过此过程可以使葡萄糖彻底降解、氧化成CO2。")
69
四. 糖酵解产物的去路 (1)在无氧或相对缺氧时 酒精发酵中:作为 乙醛 → 乙醇 的供氢体 乳酸发酵中:作为 丙酮酸 → 乳酸 的供氢体
2. NADH的去路 四. 糖酵解产物的去路 (1)在无氧或相对缺氧时 酒精发酵中:作为 乙醛 → 乙醇 的供氢体 乳酸发酵中:作为 丙酮酸 → 乳酸 的供氢体 ∴ 1分子葡萄糖通过无氧酵解,只能生成 2 个ATP
在无氧或相对缺氧时. 酒精发酵中:作为 乙醛 → 乙醇 的供氢体. 乳酸发酵中:作为 丙酮酸 → 乳酸 的供氢体. ∴ 1分子葡萄糖通过无氧酵解,只能生成. 2 个ATP.")
70
原核生物中:1分子的NADH通过呼吸链可产生3个ATP,
四. 糖酵解产物的去路 (2)在有氧条件下 原核生物中:1分子的NADH通过呼吸链可产生3个ATP, ∴ 1分子葡萄糖通过有氧酵解,可生成 2 + 3×2 = 8 个ATP 真核生物中:在植物细胞或动物的肌细胞中,1分子 的NADH通过呼吸链可产生2个ATP。 ∴ 1分子葡萄糖通过有氧酵解,可生成 2 + 2×2 = 6 个ATP
在有氧条件下. 原核生物中:1分子的NADH通过呼吸链可产生3个ATP, ∴ 1分子葡萄糖通过有氧酵解,可生成 2 + 3×2 = 8 个ATP. 真核生物中:在植物细胞或动物的肌细胞中,1分子 的NADH通过呼吸链可产生2个ATP。 ∴ 1分子葡萄糖通过有氧酵解,可生成 2 + 2×2 = 6 个ATP.")
71
如丙酮酸可转变为氨基酸,磷酸二羟丙酮可合成甘油。
五. 糖酵解的生物学意义 1. 为生物体提供一定的能量 ; 2. 糖酵解的中间物为生物合成提供原料; 如丙酮酸可转变为氨基酸,磷酸二羟丙酮可合成甘油。 3. 为糖异生作用提供了基本途径。

72
六. 糖酵解的调控 在代谢途径中,发生不可逆反应的地方常常是整个途径的调控部位,而催化这些反应的酶常常要受到调控,从而影响这些地方的反应速度,进而影响整个途径的进程。这些酶称该途径的关键酶。 在糖酵解中,有三种酶催化的不可逆反应 —— 己糖激酶、PFK(磷酸果糖激酶)、丙酮酸激酶。所以它们是关键酶。 这三种酶都是变构酶。
、丙酮酸激酶。所以它们是关键酶。 这三种酶都是变构酶。")
73
Chapter 3 The pyruvate oxidization and
第三节 丙酮酸氧化脱羧与三羧酸循环 Chapter 3 The pyruvate oxidization and citric acid cycle

74
一、丙酮酸氧化脱羧 在有氧条件下,丙酮酸氧化脱羧形成乙酰CoA,后者可进入三羧酸循环彻底氧化。 丙酮酸的氧化脱羧的部位:线粒体

75
一、丙酮酸氧化脱羧

76
催化此过程的是丙酮酸脱氢酶复合体,它由3种酶有机地组合在一起:
一、丙酮酸氧化脱羧 催化此过程的是丙酮酸脱氢酶复合体,它由3种酶有机地组合在一起: E1 —— 丙酮酸脱氢酶(pyruvate dehydrogenase PDH)。催化丙酮酸的脱羧及脱氢,形成二碳单位乙酰基。具有辅基TPP。 E2 —— 二氢硫辛酸转乙酰基酶(dihydrolipoyl transacetylase TA)。催化二碳单位乙酰基的转移。具有辅基硫辛酸。 E3 —— 二氢硫辛酸脱氢酶(dihydrolipoyl dehydrogenase DLD)。催化还原型硫辛酸→氧化型。具有辅基FAD。
。催化丙酮酸的脱羧及脱氢,形成二碳单位乙酰基。具有辅基TPP。 E2 —— 二氢硫辛酸转乙酰基酶(dihydrolipoyl transacetylase TA)。催化二碳单位乙酰基的转移。具有辅基硫辛酸。 E3 —— 二氢硫辛酸脱氢酶(dihydrolipoyl dehydrogenase DLD)。催化还原型硫辛酸→氧化型。具有辅基FAD。")
77
整个过程涉及到的6个辅因子:TPP(焦磷酸硫胺素)、SSL(硫辛酸)、FAD、NAD+、CoA、Mg2+等。
一、丙酮酸氧化脱羧 整个过程涉及到的6个辅因子:TPP(焦磷酸硫胺素)、SSL(硫辛酸)、FAD、NAD+、CoA、Mg2+等。 丙酮酸脱氢酶复合体呈圆球形,每个复合体含有: 6个PDH丙酮酸脱氢酶、24个TA氢硫辛酸转乙酰基酶、6个DLD二氢硫辛酸脱氢酶 其中TA为复合物的核心,它的一条硫辛酸臂可以旋转。
、SSL(硫辛酸)、FAD、NAD+、CoA、Mg2+等。 丙酮酸脱氢酶复合体呈圆球形,每个复合体含有: 6个PDH丙酮酸脱氢酶、24个TA氢硫辛酸转乙酰基酶、6个DLD二氢硫辛酸脱氢酶. 其中TA为复合物的核心,它的一条硫辛酸臂可以旋转。")
78
一、丙酮酸氧化脱羧

79
该循环的提出的主要贡献者是英国生化学家Krebs,所以又称Krebs循环。
1. 化学反应过程 二、TCA循环 三羧酸循环(tricarboxylic acid cycle),又叫做TCA循环,是由于该循环的第一个产物是柠檬酸,它含有三个羧基,故此得名。 该循环的提出的主要贡献者是英国生化学家Krebs,所以又称Krebs循环。 该循环还叫做柠檬酸循环。
,又叫做TCA循环,是由于该循环的第一个产物是柠檬酸,它含有三个羧基,故此得名。 该循环的提出的主要贡献者是英国生化学家Krebs,所以又称Krebs循环。 该循环还叫做柠檬酸循环。")
81
1. 化学反应过程 二、TCA循环 Step 1.乙酰CoA与草酰乙酸缩合成柠檬酸

82
The methyl carbon of acety-CoA joins the carbonyl
An aldol condensation The methyl carbon of acety-CoA joins the carbonyl carbon of oxaloacetate; citroyl-CoA is a transient intermediate; hydrolysis of the thioester bond releases a large amount of free energy.

83
Citrate synthase. Citrate is shown in green and CoA pink
1. 化学反应过程 二、TCA循环 Step 1.乙酰CoA与草酰乙酸缩合成柠檬酸 这步反应由 C4 → C6 。 Citrate synthase. Citrate is shown in green and CoA pink

84
二、TCA循环 这步反应由 C4 → C6 。 Step 1. 乙酰CoA与草酰乙酸缩合成柠檬酸
1. 化学反应过程 二、TCA循环 Step 1. 乙酰CoA与草酰乙酸缩合成柠檬酸 反应的能量由乙酰CoA的高能硫酯键提供,所以使反应不可逆。此为醇醛缩合反应,先缩合成柠檬酰CoA,然后水解。 这步反应由 C4 → C6 。

85
Iron-sulfur (red), cysteines (yellow) and isocitrate (white)
1. 化学反应过程 二、TCA循环 Step 2. 柠檬酸异构化成异柠檬酸 Iron-sulfur (red), cysteines (yellow) and isocitrate (white)
, cysteines (yellow) and isocitrate (white)")
86
1. 化学反应过程 二、TCA循环 Step 3. 异柠檬酸氧化脱羧

87
这阶段放出了1分子CO2,由 C6 → C5 ;产生1分子NADH
1. 化学反应过程 二、TCA循环 Step 3. 异柠檬酸氧化脱羧 这阶段放出了1分子CO2,由 C6 → C5 ;产生1分子NADH NADP+(gold); Ca2+(red))
; Ca2+(red))")
88
二、TCA循环 Step 4. a-酮戊二酸氧化脱羧 a-酮戊二酸脱氢酶复合体与丙酮酸脱氢酶复合体非常相似,也包含三种酶、五六种辅因子。
1. 化学反应过程 二、TCA循环 Step 4. a-酮戊二酸氧化脱羧 a-酮戊二酸脱氢酶复合体与丙酮酸脱氢酶复合体非常相似,也包含三种酶、五六种辅因子。

89
二、TCA循环 Step 4. a-酮戊二酸氧化脱羧 这阶段又放出了1分子CO2,由 C5 → C4 ;
1. 化学反应过程 二、TCA循环 Step 4. a-酮戊二酸氧化脱羧 这阶段又放出了1分子CO2,由 C5 → C4 ; 又产生1分子NADH;形成1个高能硫酯键。

90
Malonate (丙二酸) is a strong competitive inhibitor
1. 化学反应过程 二、TCA循环 Step 5. 由琥珀酰CoA生成高能磷酸键 Malonate (丙二酸) is a strong competitive inhibitor 这阶段合成了1分子高能磷酸化合物GTP
 is a strong competitive inhibitor. 这阶段合成了1分子高能磷酸化合物GTP.")
91
二、TCA循环 Step 6. 琥珀酸氧化成延胡索酸 这一阶段的反应为C4的变化; 产生1分子FADH2、1分子NADH。
1. 化学反应过程 二、TCA循环 Step 6. 琥珀酸氧化成延胡索酸 这一阶段的反应为C4的变化; 产生1分子FADH2、1分子NADH。

92
二、TCA循环 Step 7.延胡索酸至苹果酸 这一阶段的反应为C4的变化; 产生1分子FADH2、1分子NADH。
1. 化学反应过程 二、TCA循环 Step 7.延胡索酸至苹果酸 这阶段需要经历三步反应 —— 脱氢、加水、脱氢 这一阶段的反应为C4的变化; 产生1分子FADH2、1分子NADH。

93
Oxaloacetate is regenerated!
1. 化学反应过程 二、TCA循环 Step 8. 苹果酸至草酰乙酸(再生) Oxaloacetate is regenerated!
 Oxaloacetate is regenerated!")
94
二、TCA循环 Step 8. 苹果酸至草酰乙酸(再生)
1. 化学反应过程 二、TCA循环 Step 8. 苹果酸至草酰乙酸(再生) The active site of malate dehydrogenase. Malate is shown in red; NAD+ blue.
 The active site of malate dehydrogenase. Malate is shown in red; NAD+ blue.")
95
2. TCA循环的总反应 二、TCA循环

96
有2个碳原子通过乙酰CoA进入循环,以后有2个碳原子通过脱羧反应离开循环。
2. TCA循环的总反应 二、TCA循环 每经历一次TCA循环 有2个碳原子通过乙酰CoA进入循环,以后有2个碳原子通过脱羧反应离开循环。 有4对氢原子通过脱氢反应离开循环,其中3对由NADH携带,1对由FADH2携带。 产生1分子高能磷酸化合物GTP,通过它可生成1分子ATP。 消耗2分子水,分别用于合成柠檬酸(水解柠檬酰CoA)和延胡索酸的加水。
和延胡索酸的加水。")
97
二、TCA循环 产物NADH和FADH2的去路:
由TCA循环产生的NADH和FADH2必须经呼吸链将电子交给O2,才能回复成氧化态,再去接受TCA循环脱下的氢。 所以,TCA循环需要在有氧的条件下进行。否则NADH和FADH2携带的H无法交给氧,即呼吸链氧化磷酸化无法进行,NAD+及FAD不能被再生,使TCA循环中的脱氢反应因缺乏氢的受体而无法进行。

98
二、TCA循环 乙酰CoA通过TCA循环脱下的氢由NADH及FADH2经呼吸链传递给O2,由此而形成大量ATP
3.能量的化学计量 二、TCA循环 乙酰CoA通过TCA循环脱下的氢由NADH及FADH2经呼吸链传递给O2,由此而形成大量ATP 碳 源 乙酰CoA → 2CO2 能 量 1GTP → 1ATP 共12ATP 3NADH → 3ATP×3 = 9ATP 1FADH2 → 2ATP×1 = 2ATP 由乙酰CoA氧化产生的ATP中,只有1/12来自底物水平的磷酸化,其余都是由氧化磷酸化间接产生

99
二、TCA循环 碳 源 丙酮酸 → 乙酰CoA + CO2 → 3CO2 能 量 丙酮酸氧化脱羧:1NADH → 3ATP 共15ATP
3.能量的化学计量 二、TCA循环 碳 源 丙酮酸 → 乙酰CoA + CO2 → 3CO2 能 量 丙酮酸氧化脱羧:1NADH → 3ATP 共15ATP TCA循环:12ATP

100
葡萄糖有氧酵解:2ATP + 2NADH → 8 ATP
3.能量的化学计量 二、TCA循环 葡萄糖彻底氧化经由的途径: EMP途径、丙酮酸氧化脱羧、TCA循环、呼吸链氧化磷酸化。 对于原核生物: 碳 源 葡萄糖 → 2丙酮酸 → 6CO2 能 量 葡萄糖有氧酵解:2ATP + 2NADH → 8 ATP 共38ATP 丙酮酸有氧氧化:15×2 = 30 ATP

101
对于真核生物(高等植物、真菌、动物的肌细胞):
3.能量的化学计量 二、TCA循环 对于原核生物: 对于真核生物(高等植物、真菌、动物的肌细胞): 由于在EMP途径中生成的NADH在线粒体外,其磷氧比为2,所以1分子葡萄糖彻底氧化只能合成 36 ATP。
: 由于在EMP途径中生成的NADH在线粒体外,其磷氧比为2,所以1分子葡萄糖彻底氧化只能合成 36 ATP。")
102
A 柠檬酸合酶:该酶有负变构剂ATP,它使酶与底物的亲和力下降,从而Km值增大。
4. 注意点 二、TCA循环 1. 定位:线粒体 2. 不可逆反应与调节: A 柠檬酸合酶:该酶有负变构剂ATP,它使酶与底物的亲和力下降,从而Km值增大。 B 异柠檬脱氢酶:该酶有正变构剂ADP,它使酶与底物的亲和力增加。此外,NAD+、底物异柠檬酸使酶活升高;NADH、ATP使酶活下降。 C -酮二酸脱氢酶: ATP、NADH及产物琥珀酰CoA抑制酶的活性。

103
1. 为生物体提供能量,是体内主要产生ATP的途径 ;
5. TCA循环的生物学意义 二、TCA循环 1. 为生物体提供能量,是体内主要产生ATP的途径 ; 2. 循环中的中间物为生物合成提供原料; 如草酰乙酸、a-酮戊二酸可转变为氨基酸,琥珀酰CoA可用于合成叶绿素及血红素分子中的卟啉。 3. 糖类、蛋白质、脂类、核酸等代谢的枢纽。

104
1.回补反应含义: 三、TCA的回补反应 三羧酸循环的一个重要作用是它的中间物可以为生物合成提供原料,但这些中间物必须得到补充,以保证TCA循环运转。尤其是起始物草酰乙酸,缺乏它乙酰CoA就不能进入循环。 生物体中存在着及时补充草酰乙酸的反应,称为回补反应。

105
三、TCA的回补反应 丙酮酸羧化酶需要生物素作为其辅酶。 2. 回补反应的途径: 1. 丙酮酸羧化
这是动物中最重要的回补反应,在线粒体中进行。 丙酮酸羧化酶需要生物素作为其辅酶。

106
2. 回补反应的途径: 三、TCA的回补反应 2. PEP羧化酶(细胞质)
")
107
2. 回补反应的途径: 三、TCA的回补反应 3. 苹果酸酶(细胞质)
")
108
The phosphoenolpyruvate carboxykinase reaction
2. 回补反应的途径: 三、TCA的回补反应 4. PEP羧激酶(液泡) The phosphoenolpyruvate carboxykinase reaction
 The phosphoenolpyruvate carboxykinase reaction.")
109
第四节 磷酸戊糖途径

110
一、概念 HMP 途径 葡萄糖的降解除了EMP-TCA途径外,是否还存在着另外的途径? 有,什么途径? 磷酸戊糖途径。

111
一、概念 HMP途径 磷酸戊糖途径(pentose phosphate pathway or phosphogluconate pqthway),又叫做PPP,是由于该途径中有许多中间物是磷酸戊糖。 该途径又叫做磷酸葡萄糖酸途径 (hexose monophosphate shunt pathway HMP),因为磷酸葡萄糖酸是该途径的早期特征中间物。 该途径又称磷酸己糖支路,是由于从磷酸己糖开始该途径与EMP途径分支。
,因为磷酸葡萄糖酸是该途径的早期特征中间物。 该途径又称磷酸己糖支路,是由于从磷酸己糖开始该途径与EMP途径分支。")
112
二、化学历程 HMP途径 分为两个阶段: 1)糖的脱氢、脱羧:6-磷酸葡萄糖→5-磷酸核酮糖 2)糖的相互转化:6个5-磷酸核酮糖→5个6-磷酸葡萄糖
糖的脱氢、脱羧:6-磷酸葡萄糖→5-磷酸核酮糖 2)糖的相互转化:6个5-磷酸核酮糖→5个6-磷酸葡萄糖")
113
二、化学历程 HMP途径 葡萄糖氧化脱羧 第一步:脱氢 Dehydrogenation 6-P-葡萄糖 6-磷酸葡萄糖内酯

114
二、化学历程 HMP途径 6-磷酸葡萄糖内酯 G-6-P → 6-磷酸葡萄糖酸的过程不可逆. 第二步:hydrolysis 葡萄糖氧化脱羧

115
第三步:oxidative decarboxylation
二、化学历程 HMP途径 第三步:oxidative decarboxylation 葡萄糖氧化脱羧 6-P-葡萄糖酸 5-P-核酮糖 此反应也不可逆

116
Phosphopentose Isomerase
二、化学历程 HMP途径 分子重组阶段 第四步:异构化 Isomerism Phosphopentose Isomerase

117
Phosphopentose Isomerase
二、化学历程 HMP途径 第四步:异构化 Isomerism 分子重组阶段 5-P-核酮糖 5-P-核糖 Phosphopentose Isomerase

118
第五步:表异构化 Epimerization
二、化学历程 HMP途径 分子重组阶段 第五步:表异构化 Epimerization Phosphopentose Epimerase ketose xylulose

119
第五步:表异构化 Epimerization
二、化学历程 HMP途径 第五步:表异构化 Epimerization 分子重组阶段 5-P-核糖 5-P-木酮糖 Phosphopentose Epimerase

120
二、化学历程 HMP途径 第六步:转酮醇反应 Transketolase 分子重组阶段 5-P-木酮糖 5-P-核糖 3-P-甘油醛

121
第七步:转酮醇反应 Transketolase
二、化学历程 HMP途径 第七步:转酮醇反应 Transketolase 分子重组阶段 7-P-景天庚酮糖 3-P-甘油醛 4-P-赤藓糖 6-P-果糖

122
第八步:转酮醇反应 Transketolase
二、化学历程 HMP途径 第八步:转酮醇反应 Transketolase 分子重组阶段 5-P-木酮糖 4-P-赤藓糖 3-P-甘油醛 6-P-果糖

123
phosphoglucoisomerase
二、化学历程 HMP途径 第九步:异构化反应 Isomerism 分子重组阶段 磷酸已糖异构酶 phosphoglucoisomerase 6-磷酸果糖 6-磷酸葡萄糖

124
三、化学计量 HMP途径 氧化阶段 66-磷酸葡萄糖+12NADP++6H2O 65-磷酸核酮糖+6CO2+12NADPH+12H+ 非氧化重排阶段 65-磷酸核酮糖+H2O 56-磷酸葡萄糖 总反应式 6-磷酸葡萄糖+12NADP++7H2O 6CO2+12NADPH+12H++H3PO4

125
1. 产生大量的NADPH,为细胞的各种合成反应提供还原力
四、HMP途径的生物学意义 HMP途径 1. 产生大量的NADPH,为细胞的各种合成反应提供还原力 NADPH作为主要供氢体,为脂肪酸、固醇、四氢叶酸等的合成、氨的同化等反应所必需。 2. 途径中的中间物为许多化合物的合成提供原料 可以产生各种磷酸单糖。如磷酸核糖是合成核苷酸的原料,4-磷酸赤藓糖与PEP可合成莽草酸,经莽草酸途径可合成芳香族氨基酸。

126
HMP途径在生物体中普遍存在,其中动物、微生物中占糖降解的30%,植物中占50%。
3. HMP定位于细胞质,和EMP等途径相通 4. HMP在植物胁迫(如干旱、病害、伤害等)时被高速启动 HMP途径在生物体中普遍存在,其中动物、微生物中占糖降解的30%,植物中占50%。
时被高速启动. HMP途径在生物体中普遍存在,其中动物、微生物中占糖降解的30%,植物中占50%。")
127
五、磷酸戊糖途径的调控 6-磷酸葡萄糖脱氢酶为限速酶 活性受[NADP+]/[NADPH+H+]比率调节
![五、磷酸戊糖途径的调控 6-磷酸葡萄糖脱氢酶为限速酶 活性受[NADP+]/[NADPH+H+]比率调节](http://slidesplayer.com/slide/11361085/61/images/127/%E4%BA%94%E3%80%81%E7%A3%B7%E9%85%B8%E6%88%8A%E7%B3%96%E9%80%94%E5%BE%84%E7%9A%84%E8%B0%83%E6%8E%A7+6%EF%BC%8D%E7%A3%B7%E9%85%B8%E8%91%A1%E8%90%84%E7%B3%96%E8%84%B1%E6%B0%A2%E9%85%B6%E4%B8%BA%E9%99%90%E9%80%9F%E9%85%B6+%E6%B4%BB%E6%80%A7%E5%8F%97%5BNADP%2B%5D%2F%5BNADPH%2BH%2B%5D%E6%AF%94%E7%8E%87%E8%B0%83%E8%8A%82.jpg "五、磷酸戊糖途径的调控 6-磷酸葡萄糖脱氢酶为限速酶 活性受[NADP+]/[NADPH+H+]比率调节")
128
第五节 糖类的合成代谢

129
1.实验 一 光合作用 土减少了0.1千克 树重82.5千克, 增加了80千克 结论:植物生长和水有关

130
普利斯特利实验 结论:植物可以更新空气。 A 光照下,一段时间后 B C D

131
一 光合作用 生物按其利用碳源的类型可分为两类:(1) 自养生物(autotrophs),即能够吸收和利用CO2或无机碳化物,合成自身所需的全部有机物的生物。例如绿色植物、藻类、硫细菌等。这类生物利用的供氢体也是无机的,如H2O、H2S等;
 自养生物(autotrophs),即能够吸收和利用CO2或无机碳化物,合成自身所需的全部有机物的生物。例如绿色植物、藻类、硫细菌等。这类生物利用的供氢体也是无机的,如H2O、H2S等;")
132
(2) 异养生物(heterotrophs),这类生物至少需要提供一种有机物才能转变为体内所需的其它有机物的生物。例如人、动物,是利用食物中的有机物参与体内代谢而生存的。
 异养生物(heterotrophs),这类生物至少需要提供一种有机物才能转变为体内所需的其它有机物的生物。例如人、动物,是利用食物中的有机物参与体内代谢而生存的。")
133
供生命活动所需的能源主要有两种:化学能和光能。按获得能源方式不同,生物又分为两类:光养生物(phototrophs)和化养生物(chemotrophs)。绿色植物和光合细菌属于光养生物,它们在阳光的照耀下,利用光能进行着体内的代谢而茁壮成长。人类和动物属于化养生物,它们无法直接吸收光能来供机体的生命活动,只能利用有机物分解代谢产生的化学能。生物按利用营养所需的能源和碳源可分成四大类。
和化养生物(chemotrophs)。绿色植物和光合细菌属于光养生物,它们在阳光的照耀下,利用光能进行着体内的代谢而茁壮成长。人类和动物属于化养生物,它们无法直接吸收光能来供机体的生命活动,只能利用有机物分解代谢产生的化学能。生物按利用营养所需的能源和碳源可分成四大类。")
134
1 光合作用概述 光合作用(photosynthesis)是指自然界中绿色植物或光合细菌捕获光能并将其转变为化学能;再将CO2和H2O等转变为有机化合物,释放O2或S等物质的过程。1941年Cornelis Van Niel 比较了细菌和绿色植物的光合作用的特点,并用同位素18O的实验证实了光合作用可以用一个通式表示:
是指自然界中绿色植物或光合细菌捕获光能并将其转变为化学能;再将CO2和H2O等转变为有机化合物,释放O2或S等物质的过程。1941年Cornelis Van Niel 比较了细菌和绿色植物的光合作用的特点,并用同位素18O的实验证实了光合作用可以用一个通式表示:")
135
式中,H2D代表各种还原剂,是氢和电子供体。当H2D为H2O时,2D为O2,这是绿色植物将CO2还原成糖类并产生O2的机制。总反应可写成:

136
光合作用可分为两个阶段:第一阶段是光反应(light reaction),第二阶段是暗反应(dark reaction)。光反应是指光合色素将光能转变为化学能ATP和还原为NADPH的过程。暗反应即酶促反应,是指利用ATP和NADPH,将CO2还原成糖类等有机物的代谢过程。光合作用的两个阶段是偶联的关系。
,第二阶段是暗反应(dark reaction)。光反应是指光合色素将光能转变为化学能ATP和还原为NADPH的过程。暗反应即酶促反应,是指利用ATP和NADPH,将CO2还原成糖类等有机物的代谢过程。光合作用的两个阶段是偶联的关系。")
137
光能捕捉和CO2固定

138
光合作用机构

139
叶绿体结构 外 膜 内 膜 基质 基粒 基粒囊状结构的薄膜上 叶绿体中的色素分布于 。 基粒上、基质中 与光合作用有关的酶分布于 。

140
叶绿体中的色素 类胡萝卜素 叶 绿 素 (含量约占总量的3/4) 胡萝卜素 最少 (橙黄色) 叶黄素 (黄色) 较少 叶绿素a 最多
叶 绿 素 胡萝卜素 叶黄素 (橙黄色) 最少 (黄色) 较少 (含量约占总量的1/4) 叶绿素a 叶绿素 b (蓝绿色) 最多 较多 (黄绿色) (含量约占总量的3/4)
 最少. (黄色) 较少. (含量约占总量的1/4) 叶绿素a. 叶绿素 b. (蓝绿色) 最多. 较多. (黄绿色) (含量约占总量的3/4)")
141
思考:为什么春夏两季植物的叶子呈绿色,而深秋树叶则黄色呢?
由于叶绿素含量约占总量的四分之三,而类胡萝卜素仅占四分之一,叶绿素掩盖了类胡萝卜素颜色,所以通常植物的叶子总是呈绿色的。 秋天叶绿素会因为气温下降等因素的影响而分解消失;胡萝卜素和叶黄素则比较稳定,所以深秋树叶则黄色。

142
叶绿体中的类囊体薄膜上色素的分类: 天线色素 种类 绝大多数叶绿素a及全部叶绿素b、胡萝卜素和叶黄素 作用 吸收、传递光能 作用中心色素 种类 少数处于特殊状态的叶绿素a 作用 吸收、转换光能

143
光合作用意义 1.为人类活动提供能量:地球上的植物捕获太阳能,并转换为化学能贮存在有机物之中,成为我们今天工农业生产和日常生活中的重要动力资源。 2.为人类提供有机物质:绿色植物通过光合作用合成大量的有机物,对生物界的贡献是巨大的。这些有机物不仅是植物组织结构成分,也是植物各种生理活动的基础,还是人类和动物赖以生存的食物和原料。

144
光合作用意义 3.改善人类生活环境:人类和动物时刻都需要呼吸,吸入O2,吐出CO2。绿色植物光合作用时,固定CO2, 放出O2,维持了大气中的O2比例。甚至一部分O2转变为臭氧(O3),在大气中形成一种屏障,阻挡强紫外线的辐射,保护地球上的生物。另外,植树造林还可阻挡沙尘,可起到净化空气的作用。
,在大气中形成一种屏障,阻挡强紫外线的辐射,保护地球上的生物。另外,植树造林还可阻挡沙尘,可起到净化空气的作用。")
145
反应过程依据是否需要光能将光合作用分为光反应和暗反应两个阶段.

146
2H2O O2 2C3 CO2 4[H] C5 ATP 多种酶 酶 ADP+Pi C6H12O6+H2O 可见光 光反应 暗反应 基质中
片层结构 2H2O O2 2C3 CO2 固定 光解 4[H] 可见光 吸收 色素分子 还原 C5 ATP 多种酶 酶 能 ADP+Pi C6H12O6+H2O 光反应 暗反应
![2H2O O2 2C3 CO2 4[H] C5 ATP 多种酶 酶 ADP+Pi C6H12O6+H2O 可见光 光反应 暗反应 基质中](http://slidesplayer.com/slide/11361085/61/images/146/2H2O+O2+2C3+CO2+4%5BH%5D+C5+ATP+%E5%A4%9A%E7%A7%8D%E9%85%B6+%E9%85%B6+ADP%2BPi+C6H12O6%2BH2O+%E5%8F%AF%E8%A7%81%E5%85%89+%E5%85%89%E5%8F%8D%E5%BA%94+%E6%9A%97%E5%8F%8D%E5%BA%94+%E5%9F%BA%E8%B4%A8%E4%B8%AD.jpg "片层结构. 2H2O. O2. 2C3. CO2. 固定. 光解. 4[H] 可见光. 吸收. 色素分子. 还原. C5. ATP. 多种酶. 酶. 能. ADP+Pi. C6H12O6+H2O. 光反应. 暗反应.")
147
光合色素因生物不同有差异,大体分为三类: (1)叶绿素类(chlorophyll,Chl):
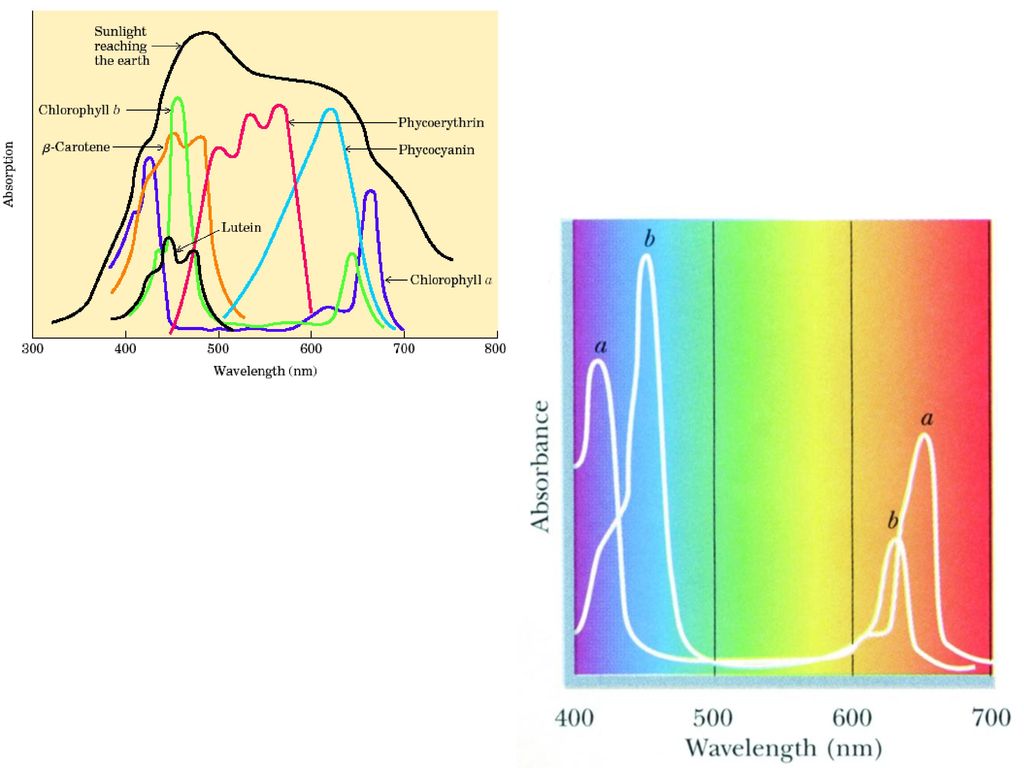
2 光反应 2.1 光合色素 光合色素因生物不同有差异,大体分为三类: (1)叶绿素类(chlorophyll,Chl): 高等植物的叶绿体中往往同时含有叶绿素a和b。这两种光合色素均是绿色的。它们的吸收光谱都在350~650 nm的可见光范围内,均有两个光吸收峰,只是峰值略有差异。如菠菜叶绿素a的两个光吸收峰分别是1=428 nm, 2=662 nm。叶绿素b的两个光吸收峰则分别是1'=452 nm, 2'=644 nm。
叶绿素类(chlorophyll,Chl): 高等植物的叶绿体中往往同时含有叶绿素a和b。这两种光合色素均是绿色的。它们的吸收光谱都在350~650 nm的可见光范围内,均有两个光吸收峰,只是峰值略有差异。如菠菜叶绿素a的两个光吸收峰分别是1=428 nm, 2=662 nm。叶绿素b的两个光吸收峰则分别是1 =452 nm, 2 =644 nm。")
149
(2) 类胡萝卜素(carotenoid) 类胡萝卜素存在于所有植物、藻类、光合细菌的光合细胞之中。作为辅助色素的类胡萝卜素已发现有70多种。其中最重要的是橙红色的-胡萝卜素和黄色的叶黄素(lutein)。类胡萝卜素的吸收光谱在400~500 nm的可见光范围内。它们吸收光能并将光能传递给叶绿素a,再由叶绿素将光能传递给光合作用中心,发生光化学反应。
。类胡萝卜素的吸收光谱在400~500 nm的可见光范围内。它们吸收光能并将光能传递给叶绿素a,再由叶绿素将光能传递给光合作用中心,发生光化学反应。")
150
(3) 藻胆色素 藻胆色素类主要有藻蓝素(phycocyanobilin)和藻红素(phycoerythrobilin),存在于藻类和某些细菌中并能够与蛋白质结合的一类色素。与这些色素结合的蛋白质称为藻胆蛋白。藻胆色素类分子中主要含有开环四吡咯结构,吸收光谱在400~500 nm的可见光范围内。它们也是辅助色素,能够吸收光能并将光能完全传递给叶绿素a。数百万个藻胆蛋白高度有序地缔合在一起的集装体称为藻胆体(phycobilisome)。它们是藻类生物的主要集光器。
和藻红素(phycoerythrobilin),存在于藻类和某些细菌中并能够与蛋白质结合的一类色素。与这些色素结合的蛋白质称为藻胆蛋白。藻胆色素类分子中主要含有开环四吡咯结构,吸收光谱在400~500 nm的可见光范围内。它们也是辅助色素,能够吸收光能并将光能完全传递给叶绿素a。数百万个藻胆蛋白高度有序地缔合在一起的集装体称为藻胆体(phycobilisome)。它们是藻类生物的主要集光器。")
152
2.2 光反应系统 (1) 光合单位(photosynthetic unit) 一个光合单位包括几百个集光叶绿素或其它色素分子,收集入射光,通过诱导共振方式进行能量的传递单元。
 光合单位(photosynthetic unit) 一个光合单位包括几百个集光叶绿素或其它色素分子,收集入射光,通过诱导共振方式进行能量的传递单元。")
153
(2) 反应中心色素(reaction center pigment)
反应中心色素是一个具有光化学反应特性的叶绿素a“特殊对”(二聚体)。中心色素能接受其它叶绿素等光合色素的能量。当中心色素的叶绿素分子被氧化时,就变为一个阳离子自由基Chla.+ ,Chla.+作为电子受体,参与光化学反应。
。中心色素能接受其它叶绿素等光合色素的能量。当中心色素的叶绿素分子被氧化时,就变为一个阳离子自由基Chla.+ ,Chla.+作为电子受体,参与光化学反应。")
154
(3) 光系统(photosystem) 光系统是一个完整的具对光的吸收、能量传递和转换,由多种色素和蛋白质组成的独立结构。光系统分为光系统I(photosystem I,PSI)和光系统II(photosystem II,PSII)。
 光系统(photosystem) 光系统是一个完整的具对光的吸收、能量传递和转换,由多种色素和蛋白质组成的独立结构。光系统分为光系统I(photosystem I,PSI)和光系统II(photosystem II,PSII)。")
155
PSI于1955年由Duysens发现的。PSI中心色素分子的吸收峰值为700 nm,可用代号P700表示,中心色素复合蛋白属于铁氧还蛋白型,类似绿色硫细菌的光合系统。
PSII是1956年由Kok B发现的。它的中心色素分子吸收峰值为680 nm,可用代号P680表示,中心色素复合蛋白属于脱镁叶绿素-醌型,类似紫色细菌的光系统。

156
2.3 光反应 光反应的最初阶段是色素分子吸收光能进行氧化还原反应,以及光能转变为电能的过程。当聚光色素分子吸收光能并将其传递给作用中心时,作用中心色素分子(P)由基态变为激发态(P*),一个电子跃迁到较高的外轨道上。这种激发态极不稳定,只能维持10-11~10-9秒。当激发态P*恢复到基态时,释放的多余能量,可转变成多种形式,
由基态变为激发态(P*),一个电子跃迁到较高的外轨道上。这种激发态极不稳定,只能维持10-11~10-9秒。当激发态P*恢复到基态时,释放的多余能量,可转变成多种形式,")
158
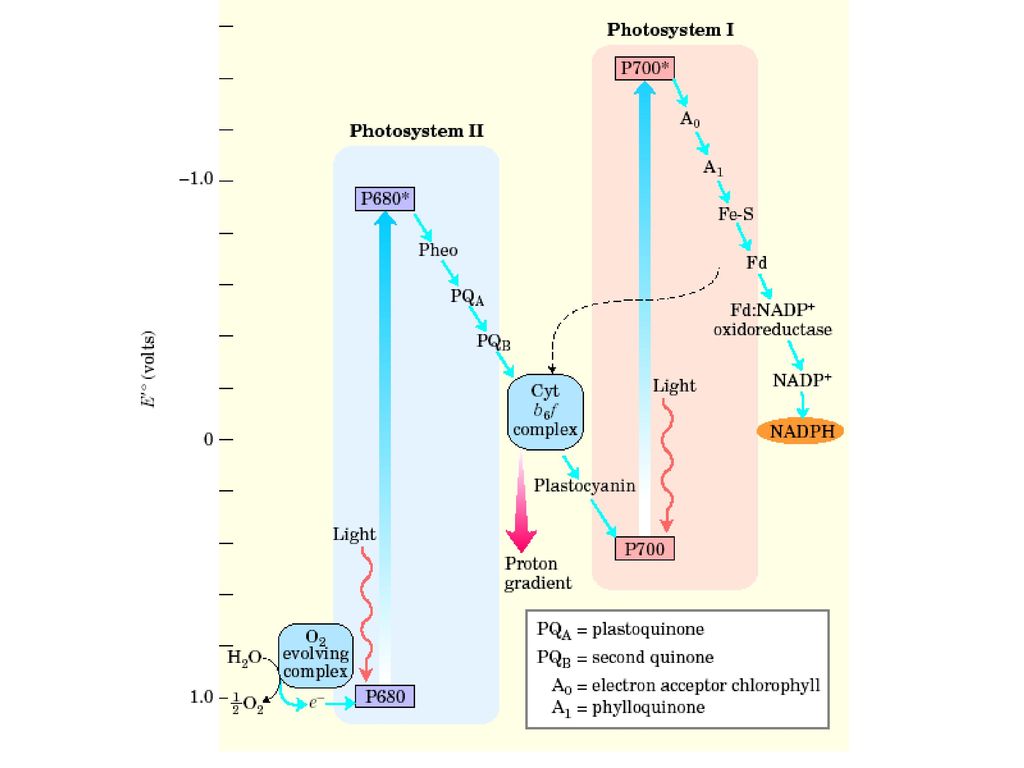
2.4光合电子传递链(Z图式) 光合电子传递链,简称光合链(photosynthetic chain),主要指以植物为代表的叶绿体类囊体膜上有序地排列着电子传递体,两个光系统串联在其中。光驱动电子从H2O流向NADP+,H2O被氧化、光解,释放出O2;NADP+变为具有还原力的NADPH。质子跨膜梯度为光能转变为化学能提供了条件。光合链图形象侧着排的Z字母,所以也叫Z图式(Z scheme)。
,主要指以植物为代表的叶绿体类囊体膜上有序地排列着电子传递体,两个光系统串联在其中。光驱动电子从H2O流向NADP+,H2O被氧化、光解,释放出O2;NADP+变为具有还原力的NADPH。质子跨膜梯度为光能转变为化学能提供了条件。光合链图形象侧着排的Z字母,所以也叫Z图式(Z scheme)。")
159
光合电子传递链(Z图式)
")
161
光合电子传递链成员 PQ—质体醌;Cytb6f—细胞色素b6、细胞色素f复合体;PC—质蓝素;A0—特殊叶绿素a; A1—叶绿醌;FeS—铁硫蛋白;Fd—铁氧还蛋白;FP—Fd-NADP氧化还原酶

162
光合电子传递链成员 PQ—质体醌;Cytb6f—细胞色素b6、细胞色素f复合体;PC—质蓝素;A0—特殊叶绿素a; A1—叶绿醌;FeS—铁硫蛋白;Fd—铁氧还蛋白;FP—Fd-NADP氧化还原酶

163
2.5 光合磷酸化 光合磷酸化(photophosphorylation)光合生物细胞利用光能驱动光合链的电子传递,引起质子形成跨膜梯度和电位差,膜上CF0CF1-ATP合酶利用质子返回势能,使ADP磷酸化形成ATP的过程。
光合生物细胞利用光能驱动光合链的电子传递,引起质子形成跨膜梯度和电位差,膜上CF0CF1-ATP合酶利用质子返回势能,使ADP磷酸化形成ATP的过程。")
164
2.5 光合磷酸化

165
ADP+Pi ATP 2H2O O2 酶 光反应阶段 条件 : 场所: 物质变化: 能量变化: 叶绿体内的类囊体膜上 光、 色素、 酶
色素分子 ADP+Pi ATP 2H2O O2 酶 吸收 光解 能 可见光 光反应 光反应阶段 条件 : 场所: 物质变化: 能量变化: 叶绿体内的类囊体膜上 光、 色素、 酶 H2O [H] + O2 光能 水的光解: (还原剂) ADP+Pi +能量(光能) ATP 酶 ATP的合成: 光能转变为活跃的化学能贮存在ATP中
 ADP+Pi +能量(光能) ATP. 酶. ATP的合成: 光能转变为活跃的化学能贮存在ATP中.")
166
二 二氧化碳固定

167
2C3 [H] CO2 ATP C5 酶 多种酶 ADP+Pi C6H12O6+H2O 暗反应阶段 条件: 场所: 物质变化: 能量变化:
叶绿体的基质中 多种酶、 [H] 、ATP CO2+C C3 酶 CO2的固定: 2C3 酶 糖类 (CH2O) C3的还原: ADP+Pi ATP [H] 、 ATP中活跃的化学能转变为糖类等 有机物中稳定的化学能 C5 2C3 多种酶 固定 还原 C6H12O6+H2O CO2 ADP+Pi ATP 酶 能 [H] 暗反应 暗反应
![2C3 [H] CO2 ATP C5 酶 多种酶 ADP+Pi C6H12O6+H2O 暗反应阶段 条件: 场所: 物质变化: 能量变化:](http://slidesplayer.com/slide/11361085/61/images/167/2C3+%5BH%5D+CO2+ATP+C5+%E9%85%B6+%E5%A4%9A%E7%A7%8D%E9%85%B6+ADP%2BPi+C6H12O6%2BH2O+%E6%9A%97%E5%8F%8D%E5%BA%94%E9%98%B6%E6%AE%B5+%E6%9D%A1%E4%BB%B6%EF%BC%9A+%E5%9C%BA%E6%89%80%EF%BC%9A+%E7%89%A9%E8%B4%A8%E5%8F%98%E5%8C%96%EF%BC%9A+%E8%83%BD%E9%87%8F%E5%8F%98%E5%8C%96%EF%BC%9A.jpg "叶绿体的基质中. 多种酶、 [H] 、ATP. CO2+C5 2C3. 酶. CO2的固定: 2C3. 酶. 糖类. (CH2O) C3的还原: ADP+Pi. ATP. [H] 、 ATP中活跃的化学能转变为糖类等. 有机物中稳定的化学能. C5. 2C3. 多种酶. 固定. 还原. C6H12O6+H2O. CO2. ADP+Pi. ATP. 酶. 能. [H] 暗反应. 暗反应.")
168
由于途径中的最初产物是三碳化合物(3-磷酸甘油酸),故也称作C3途径。
1945年美国的Calvin M等人利用单细胞小球藻作为实验材料,应用14C示踪技术和双向纸层析法,经过十年的研究揭示了光合作用暗反应阶段碳素同化及受体再生的循环途径,因此称为卡尔文循环(Calvin –Benson cycle)。 由于途径中的最初产物是三碳化合物(3-磷酸甘油酸),故也称作C3途径。
。 由于途径中的最初产物是三碳化合物(3-磷酸甘油酸),故也称作C3途径。")
169
1.1 CO2的固定 整个循环可分为三个阶段:(1) CO2固定
CO2的固定(CO2 fixation),即游离的CO2经酶促反应转变为有机物分子中的羧基,也称作CO2的羧化(carboxylation)。 CO2的受体是核酮糖-1,5-二磷酸(ribulose-1,5-bisphosphate,RuBP),催化此反应的酶是核酮糖-1, 5-二磷酸羧化酶/加氧酶(ribulose-1, 5-bisphosphate carboxylase / oxygenase,Rubisco)。
,即游离的CO2经酶促反应转变为有机物分子中的羧基,也称作CO2的羧化(carboxylation)。 CO2的受体是核酮糖-1,5-二磷酸(ribulose-1,5-bisphosphate,RuBP),催化此反应的酶是核酮糖-1, 5-二磷酸羧化酶/加氧酶(ribulose-1, 5-bisphosphate carboxylase / oxygenase,Rubisco)。")
170
1.1 CO2的固定 整个循环可分为三个阶段:(1) CO2固定 上述反应在夜间或黑暗条件下受阻 1, 5-二磷酸羧化酶
核酮糖-1,5-二磷酸

171
CO2的固定反应是卡尔文循环的限速步骤。其关键酶——Rubisco是个双功能的酶。它的活性中心有两种功能:既可以催化上述羧化反应,又可以催化加氧反应(在光呼吸中)。该酶位于叶绿体基质中,约占叶子可溶性蛋白的50%,是植物中含量最丰富的酶。 1, 5-二磷酸羧化酶

172
1.1 CO2的固定 Rubisco活性中心受CO2和Mg2+以及ATP的活化。酶的最适pH≈8。已知底物RuBP与Rubisco结合较牢,反而降低了羧化反应的速度,在Rubisco激活酶(Rubisco activase)的调节下,可以促进RuBP的释放,活化了Rubisco。活化过程 中Rubisco激活酶需要ATP,因ATP来自于光反应,所以光是Rubisco的间接激活剂。
的调节下,可以促进RuBP的释放,活化了Rubisco。活化过程 中Rubisco激活酶需要ATP,因ATP来自于光反应,所以光是Rubisco的间接激活剂。")
173
1.2 羧化产物的还原 这一阶段包括两步反应:(1)在磷酸甘油酸激酶催化下,发生高能键的转移,产生1,3-二磷酸甘油酸(BPG);(2)在磷酸甘油醛脱氢酶的催化下,还原产生3-磷酸甘油醛(G3P)。第二步酶是光调节酶。反应中消耗的ATP和NADPH来自于前面介绍的光合作用光反应所产生的化学能和还原力。
在磷酸甘油酸激酶催化下,发生高能键的转移,产生1,3-二磷酸甘油酸(BPG);(2)在磷酸甘油醛脱氢酶的催化下,还原产生3-磷酸甘油醛(G3P)。第二步酶是光调节酶。反应中消耗的ATP和NADPH来自于前面介绍的光合作用光反应所产生的化学能和还原力。")
174
1.3 受体的再生 在磷酸丙糖异构酶的催化下,3-磷酸甘油醛(G3P)异构为磷酸二羟丙酮(DHAP);接着在醛缩酶的催化下,缩合成果糖-1, 6-二磷酸(FBP);然后在光调节酶:果糖-1, 6-二磷酸酶(fructose -1,6-bisphosphatase, FBPase)的催化下,水解去掉一个磷酸基团,变为果糖-6-磷酸(F6P);再经磷酸葡萄糖异构酶催化,就变为葡糖-6-磷酸(G6P)
异构为磷酸二羟丙酮(DHAP);接着在醛缩酶的催化下,缩合成果糖-1, 6-二磷酸(FBP);然后在光调节酶:果糖-1, 6-二磷酸酶(fructose -1,6-bisphosphatase, FBPase)的催化下,水解去掉一个磷酸基团,变为果糖-6-磷酸(F6P);再经磷酸葡萄糖异构酶催化,就变为葡糖-6-磷酸(G6P)")
175
1.3 受体的再生 果糖-6-磷酸 葡糖-6-磷酸

176
1.3 受体的再生 磷酸丙糖是代谢中的一个转折点,除了参与卡尔文循环中的反应外,还有其它去路:① 转化为淀粉,作为植物贮存形式备用;② 外运到细胞质中先形成蔗糖,再转移到植物生长的区域;③ 在发育的叶片中,相当一部分磷酸丙糖转移到细胞质,进入糖酵解、TCA循环等途径为生物供能。

177
1.3 受体的再生 5-磷酸木酮糖

178
1.3 受体的再生 醛缩酶还可将上述反应产生的4-磷酸赤藓糖与磷酸二羟丙酮缩合,形成七碳糖:景天庚酮糖-1, 7-二磷酸(SBP)。在景天庚酮糖二磷酸酶(sedoheptulose bisphosphatase,SBPase)的作用下,SBP被水解,去除一个磷酰基,成为景天庚酮糖-7-磷酸(S7P)。此水解酶是光调节酶,也是植物中特有的酶
。在景天庚酮糖二磷酸酶(sedoheptulose bisphosphatase,SBPase)的作用下,SBP被水解,去除一个磷酰基,成为景天庚酮糖-7-磷酸(S7P)。此水解酶是光调节酶,也是植物中特有的酶.")
179
4-磷酸赤藓糖 磷酸二羟丙酮 景天庚酮糖-7-磷酸 景天庚酮糖-1, 7-二磷酸
1.3 受体的再生 4-磷酸赤藓糖 磷酸二羟丙酮 景天庚酮糖-7-磷酸 景天庚酮糖-1, 7-二磷酸

180
1.3 受体的再生 转羟乙醛酶还可以催化酮糖供体“景天庚酮糖-7-磷酸”将羟乙醛基转移给醛糖受体“3-磷酸甘油醛”,产生5-磷酸核糖(R5P)和5-磷酸木酮糖(Xu5P)。
和5-磷酸木酮糖(Xu5P)。")
181
1.3 受体的再生 再经两种异构酶催化,可发生五碳糖的转变,其共同的目标是形成5-磷酸核酮糖(Ru5P)。Xu5P经磷酸核酮糖差向异构酶催化,C3上羟基位置的改变,就可以变成Ru5P 。 5-磷酸核酮糖 5-磷酸木酮糖

182
1.3 受体的再生 R5P经磷酸核糖异构酶催化,醛糖就可以变成酮糖Ru5P 5-磷酸核糖 5-磷酸核酮糖

183
1.3 受体的再生 最后在磷酸核酮糖激酶(phosphoribulose kinase)催化下, 消耗1ATP, 产生1, 5-二磷酸核酮糖(RuBP),完成了受体的再生。
催化下, 消耗1ATP, 产生1, 5-二磷酸核酮糖(RuBP),完成了受体的再生。")
184
⑴ Rubisco*; ⑵ 磷酸甘油酸激酶; ⑶ 磷酸甘油醛脱氢酶* ⑷ 磷酸丙糖异构酶; ⑸ 醛缩酶; ⑹ FBPase*; ⑺ 磷酸葡糖异构酶; ⑻ 磷酸葡糖酶; ⑼ 转羟乙醛酶; ⑽ 醛缩酶; ⑾ SBPase*; ⑿ 转羟乙醛酶; ⒀ 磷酸核酮糖差向异构酶; ⒁ 磷酸核糖异构酶; ⒂ 磷酸核酮糖激酶*

185
1.3 受体的再生 总反应式 或 光合作用的卡尔文循环中,每同化一分子CO2,需要消耗三分子ATP和二分子的NADPH。

186
2 C4 途径 C4途径是一种在光合作用中与卡尔文循环有一定联系的辅助途径。该途径的作用是固定、浓缩和转运CO2到C3途径所在的维管束鞘细胞中,使其中的CO2浓度升高,提高其光合作用速率。因CO2固定最初产物是四碳二羧酸,固称为C4途径。通过C4途径固定、同化CO2的植物,简称为C4植物,如玉米、甘蔗、高粱、苋菜、狼尾草、黍等。

187
2.1 C4途径与C3途径的关系 C3植物与C4植物光合作用的差异在于暗反应阶段:C3植物的CO2固定及碳素同化途径均在叶肉细胞的叶绿体中进行。C4植物的CO2固定先在靠近叶表面的叶肉细胞中进行,然后经C4途径的四碳二羧酸的转移,把CO2运输到叶片内部维管束鞘细胞的叶绿体中,再进行卡尔文循环。可见C4植物碳同化过程应包括C4途径和C3途径。

188
2.1 C4途径与C3途径的关系

189
C4途径将CO2和Rubisco (1, 5-二磷酸羧化酶)
从氧含量较高的叶片表面转移到氧浓度较低的内部维管束鞘细胞之中,再进行CO2的固定反应。其转移的目的是避免 Rubisco 发生另一种浪费资源的加氧反应。热带禾本科植物采用C4途径的原因之一是克服了高温条件下CO2和Rubisco的亲和力的下降。

190
C4途径CO2的最初受体是一种三碳化合物——磷酸烯醇式丙酮酸(PEP),在PEP羧化酶催化下,固定CO2,生成草酰乙酸(OAA)
,在PEP羧化酶催化下,固定CO2,生成草酰乙酸(OAA)")
191
这个反应在叶肉细胞质中进行。PEP羧化酶对CO2的亲和力很强,其Km≈7 mol/L,而此时的Rubisco的Km≈450 mol/L。
PEP羧化酶从高O2低CO2的叶肉中固定CO2,为CO2的转移、同化奠定了基础。PEP羧化酶是一种调节酶,被CO2固定的反应产物OAA所抑制,受EMP途径中的葡萄糖-6-磷酸(G6P)的活化。该酶遇到光也活化,视作光合作用的酶。
的活化。该酶遇到光也活化,视作光合作用的酶。")
192
在叶肉细胞的叶绿体中,经NADP-苹果酸脱氢酶催化,转变为苹果酸。
2.2 C4途径中主要的酶促反应 在叶肉细胞的叶绿体中,经NADP-苹果酸脱氢酶催化,转变为苹果酸。

193
在叶肉细胞质内,由谷草转氨酶催化,转变为天冬氨酸
2.2 C4途径中主要的酶促反应 在叶肉细胞质内,由谷草转氨酶催化,转变为天冬氨酸

194
上述苹果酸或天冬氨酸均能够从叶肉细胞转移到维管束鞘细胞中。这些四碳二羧酸起着CO2携带体的作用。
在维管束鞘细胞中,它们发生脱羧反应,放出CO2,以利于CO2进入卡尔文循环,再参与糖的合成。脱羧反应因植物不同;也相应有差异,主要有三种形式:

195
在维管束鞘细胞的叶绿体中,经NADP-苹果酸酶催化,氧化脱羧变为丙酮酸。属于这类的植物是玉米、甘蔗、高粱等。
2.2 C4途径中主要的酶促反应 在维管束鞘细胞的叶绿体中,经NADP-苹果酸酶催化,氧化脱羧变为丙酮酸。属于这类的植物是玉米、甘蔗、高粱等。

196
在维管束鞘线粒体中,经NAD-苹果酸酶作用,氧化脱羧,形成丙酮酸。
2.2 C4途径中主要的酶促反应 在维管束鞘线粒体中,经NAD-苹果酸酶作用,氧化脱羧,形成丙酮酸。

197
2.2 C4途径中主要的酶促反应 脱羧后,形成的丙酮酸先经转氨酶作用变为丙氨酸,从维管束鞘细胞转移到叶肉细胞的叶绿体中,再经转氨酶作用又转变回丙酮酸,然后通过丙酮酸磷酸双激酶(pyruvate phosphate dikinase)的催化才转变成PEP,完成C4途径受体的再生。
的催化才转变成PEP,完成C4途径受体的再生。")
198
3 糖异生作用 葡萄糖异生作用(gluconeogenesis)是指以非糖有机物作为前体合成为葡萄糖的过程。这是植物、动物体内一种重要的单糖合成途径。非糖物质包括乳酸、丙酮酸、甘油、草酰乙酸、乙酰CoA以及生糖氨基酸(如丙氨酸)等。 植物果实成熟期间,有机酸含量下降,糖份含量增加,就是葡糖异生作用的结果。动物机体内葡萄糖异生作用是必不可少的。它对维持血糖浓度的恒定,为大脑、肌肉、眼晶状体、中枢神经系统等组织利用葡糖分解供能提供了保障。

199
3.1 葡萄糖异生途径 葡萄糖异生途径几乎是EMP途径的逆转,但要绕过EMP途径三处不可逆反应,采用葡萄糖异生作用特有的酶催化、转移,才能完成非糖有机物合成为葡萄糖的过程。葡萄糖异生途径三处迂回路径是:

200
3.1 葡萄糖异生途径 (1) 丙酮酸磷酸烯醇式丙酮酸
 丙酮酸磷酸烯醇式丙酮酸")
201
3.1 葡萄糖异生途径 (2) 果糖 –1, 6 -二磷酸 果糖- 6-磷酸 二磷酸果糖脂酶
 果糖 –1, 6 -二磷酸 果糖- 6-磷酸 二磷酸果糖脂酶")
202
3.1 葡萄糖异生途径 (3) 葡萄糖-6-磷酸 葡萄糖 6-磷酸葡萄糖脂酶
 葡萄糖-6-磷酸 葡萄糖 6-磷酸葡萄糖脂酶")
203
4 蔗糖和多糖的生物合成 4.1 糖核苷酸的作用 在高等植物、动物体内,游离的单糖不能参与双糖和多糖的合成反应,延长反应中提供的单糖基必须是活化的糖供体,这种活化的糖是一类糖核苷酸,即糖与核苷酸结合的化合物。糖核苷酸的作用是作为双糖或多糖,甚至是糖蛋白等复合糖合成过程中参与延长单糖基的活化形式或供体。最早发现的糖核苷酸是尿苷二磷酸葡萄糖(uridine diphosphate glucose, UDPG)
")
204
UDPG结构式

205
UDPG 合成反应

206
4.2 蔗糖的生物合成 (1) 磷酸蔗糖合酶途径 该途径存在于光合组织的细胞质中。被认为是植物合成蔗糖的主要途径。磷酸蔗糖合酶(sucrose phosphate synthase)属于转移酶类。它利用UDPG作为葡萄糖的供体,以F6P为葡萄糖的受体,反应产物是蔗糖-6-磷酸;再通过磷酸蔗糖酶水解,脱去磷酸基团而生成蔗糖。
属于转移酶类。它利用UDPG作为葡萄糖的供体,以F6P为葡萄糖的受体,反应产物是蔗糖-6-磷酸;再通过磷酸蔗糖酶水解,脱去磷酸基团而生成蔗糖。")
207
(1) 磷酸蔗糖合酶途径 果糖- 6-磷酸
 磷酸蔗糖合酶途径 果糖- 6-磷酸")
208
在非光合组织中蔗糖合酶(sucrose synthase)活性较高。蔗糖合酶也属于转移酶类,可催化糖基转移。
4.2 蔗糖的生物合成 (2) 蔗糖合酶途径 在非光合组织中蔗糖合酶(sucrose synthase)活性较高。蔗糖合酶也属于转移酶类,可催化糖基转移。
 蔗糖合酶途径. 在非光合组织中蔗糖合酶(sucrose synthase)活性较高。蔗糖合酶也属于转移酶类,可催化糖基转移。")
209
式中;①引物的n≥3;②活化的葡糖基从引物的非还原端延长;③淀粉合酶催化连接的键是-1,4糖苷键。
4.3 淀粉和糖原的生物合成 4.3.1 直链淀粉的生物合成 ① 淀粉合酶(starch synthase) 式中;①引物的n≥3;②活化的葡糖基从引物的非还原端延长;③淀粉合酶催化连接的键是-1,4糖苷键。
 式中;①引物的n≥3;②活化的葡糖基从引物的非还原端延长;③淀粉合酶催化连接的键是-1,4糖苷键。")
210
10. 3.3 淀粉和糖原的生物合成 4.3.1 直链淀粉的生物合成 ② D酶
D酶是一种糖苷基转移酶,作用的键是α-1,4糖苷键;转移的基团主要是麦芽糖残基;催化的底物可以是葡萄糖、麦芽多糖;起着加成反应作用。故有人称之为加成酶。D酶的存在,有利于葡萄糖转变为麦芽多糖,为直链淀粉延长反应提供了必要的引物。

211
淀粉和糖原的生物合成 4.3.1 直链淀粉的生物合成 ③ 蔗糖转化为淀粉

212
淀粉和糖原的生物合成 4.3.1 直链淀粉的生物合成 ③ 蔗糖转化为淀粉

213
淀粉和糖原的生物合成 支链淀粉的生物合成 支链淀粉分支点α-1, 6糖苷键的形成需要淀粉分支酶又称Q酶(Q enzyme)作用,将直链淀粉的一部分拆下来,装配成侧链。Q酶具有双重功能:既能催化直链淀粉的α-1, 4糖苷键的断裂,又能催化α-1, 6糖苷键的连接,形成支链淀粉。
作用,将直链淀粉的一部分拆下来,装配成侧链。Q酶具有双重功能:既能催化直链淀粉的α-1, 4糖苷键的断裂,又能催化α-1, 6糖苷键的连接,形成支链淀粉。")
214
支链淀粉的生物合成

Similar presentations





 反应部位 器官,细胞内定位 生理意义 代谢调节 主要调节点,主要变构抑制剂、变构激活剂 各代谢途径之间的联系和调控.>")



 一、叶绿体的形态 二、结构与化学组成 三、叶绿体蛋白的定位 四、叶绿体的功能——光合作用>")
 合成代谢------需要能量 物质代谢 能量代谢>")













