Download presentation
1
第八章 糖类代谢 糖类是生物体的基本营养物质和重要组成部分,是生物体在生命活动中的主要能源和 碳源,同时糖类物质也是现代发酵工业中最常用的原料。近年来科学研究证明,糖及其糖链在生命活动中担负着极为重要的信息功能。通过研究糖代谢的规律,从而找出代谢特点和条件,为人类的发展提供所需产品。糖代谢可分为分解代谢和合成代谢两个方面,生物体内的糖代谢基本过程相类似。

2
第一节 多糖的降解 多糖是由单糖组成的大分子糖类,如淀粉、纤维素、菊糖、琼脂、半纤维素、果胶质等,大量存在于植物体内。微生物细胞中的多糖可分为两类:一类为细胞的贮藏物质,在细胞内呈不溶性颗粒;另一类组成细胞结构。

3
一、淀粉酶与淀粉的降解 a-淀粉酶 a-淀粉酶又叫淀粉-1,4-糊精酶,液化酶,系统名称a-1,4-葡聚糖葡聚糖水解酶,编号E.C 。工业酶制剂主要靠芽孢杆菌发酵生产,菌种有枯草芽孢杆菌BF7658,发酵单位可达300~400u/ml。

4
a-淀粉酶是一种内切酶,该酶能在淀粉链的内部随机切割a-1,4-葡糖苷键,生成小分子糊精及少量麦芽糖和葡萄糖,生成的麦芽糖和葡萄糖都是a-型。其特点是底物分子越大,即淀粉链越长,水解的速度越快,淀粉溶液的粘度迅速下降。当淀粉被水解成为短链糊精时,水解速度就很慢,要使糊精进一步水解则需很长时间。故该酶的主要作用是使淀粉水解生成糊精。

5
a-淀粉酶虽然能水解淀粉链的a-1,4-葡糖苷键,但它不能水解麦芽糖,它的最小作用底物是麦芽三糖。此酶不能水解a-1,6-葡糖苷键,所以它作用于支链淀粉时,有异麦芽糖产生。
a-淀粉酶的用途很广。酶的性质不同,使用的针对性有所不同,例如:米曲霉a-淀粉酶比较耐酸,可用作消化药物;霉菌a-淀粉酶耐热性较差,适用于面包制作;高温a-淀粉酶适用于原料的工业化处理。工业水解淀粉时,a-淀粉酶的用量为30~60u/g淀粉。

6
b-淀粉酶 b-淀粉酶又叫淀粉-1,4-麦芽糖苷酶,系统名称a-1,4葡聚糖麦芽糖苷酶,编号E.C 。此酶分布于植物和微生物中,现微生物生产主要有芽孢杆菌属的蜡状芽孢杆菌、巨大芽孢杆菌、凝结芽孢杆菌、多粘芽孢杆菌等。

7
b-淀粉酶是一种外切酶,作用于淀粉时,从淀粉链的非还原端开始,水解它的a-D-1,4-葡萄糖苷键,水解时沿着淀粉链每次水解掉两个葡萄糖单位,水解产物为麦芽糖。由于它只能从淀粉链的外部开始依次进行水解,故水解速度较慢,不能像a-淀粉酶那样使淀粉溶液粘度迅速降低。该酶不能水解也不能绕过a-1,6-葡萄糖苷键。当其作用于支链淀粉时,遇到分支点即停止作用,剩下的大分子量分支糊精,称为b-极(界)限糊精。b-淀粉酶水解淀粉产生的游离半缩酸羟基发生一个沃尔登(Walden—inversion)转位作用,将a-型转变为b-型,生成b-麦芽糖。
限糊精。b-淀粉酶水解淀粉产生的游离半缩酸羟基发生一个沃尔登(Walden—inversion)转位作用,将a-型转变为b-型,生成b-麦芽糖。")
8
b-淀粉酶主要用于淀粉糖的生产,如贻糖、高麦芽糖浆等;由于其麦芽糖产率高,故在啤酒工业中广为应用。

9
g-淀粉酶 g-淀粉酶又称葡萄糖淀粉酶、糖化酶、淀粉-葡萄糖苷酶,系统命名a-1,4-葡聚糖葡萄糖水解酶。编号E.C 。大量的工业用糖化酶主要利用霉菌发酵生产。

10
g-淀粉酶是一种外切酶,该酶能水解淀粉的a-1,4葡萄糖苷键和a-1,6葡萄糖苷键。其作用方式是从淀粉链的非还原端开始,依次水解它的a-1,4-葡萄糖苷键,将葡萄糖一个一个水解下来,对于支链淀粉,当水解到分支点时,一般先将a-1,6葡萄糖苷键断开,然后继续水解,所以能将支链淀粉全部水解成葡萄糖。该酶水解淀粉也能产生转位作用,所以产物为b-葡萄糖。

11
g-淀粉酶虽然能水解a-1,6葡萄糖苷键,但不能水解异麦芽糖,即不能水解单独存在的a-1,6葡萄糖苷键,但它能水解b-极限糊精。

12
异淀粉酶 异淀粉酶又称淀粉-1,6-葡萄糖苷酶,系统名称为葡聚糖a-1,6-葡聚糖水解酶,编号E.C 。也有根据其来源与作用特点称其为脱支酶、Q酶、R酶、普鲁蓝酶等。该类酶是一种内切酶,能水解支链淀粉(糖原)分子中的a-1,6-葡糖苷键,使支链淀粉变成直链状的糊精。 其工业用酶制剂主要由微生物发酵生产。在工业生产中,一般与其它酶配合使用,可降低残糖,提高出酒率。

14
糖的消化吸收 淀粉(starch) 口腔,-淀粉酶(-amylase),少量作用 胃, 几乎不作用
胃, 几乎不作用 小肠,胰-amylase,主要的消化场所 麦芽糖、糊精、蔗糖、乳糖等(食物中所混入) 麦芽糖酶,糊精酶,蔗糖酶,乳糖酶 葡萄糖、半乳糖、果糖 肠黏膜细胞肠壁毛细血管门静脉血液 组织、细胞
 麦芽糖酶,糊精酶,蔗糖酶,乳糖酶. 葡萄糖、半乳糖、果糖. 肠黏膜细胞肠壁毛细血管门静脉血液 组织、细胞.")
15
二、 纤维素酶与纤维素的降解 纤维素酶是水解纤维素的一类酶的总称。纤维素酶的组分很多,二十世纪中期,Reese等人提出了关于纤维素酶作用方式的C1-Cx假说,即纤维素酶至少包括三种类型:破坏纤维素晶状结构的C1酶,水解游离(直链)纤维素分子的Cx酶,水解纤维二糖的b-葡萄糖苷酶。
纤维素分子的Cx酶,水解纤维二糖的b-葡萄糖苷酶。")
16
纤维素酶的存在方式有两种:胞外酶,溶解游离于培养基中,霉菌中产生的纤维素酶属于这种形式;细胞表面酶,结合存在于细胞表面上,如粘细菌的纤维素酶存在于细胞壁内。
人类和高等动、植物都不能合成纤维素酶类,因而自身不能消化纤维素。反刍动物的瘤胃中生存有大量产纤维素酶微生物,所以能以纤维素作为营养物质。产生纤维素酶的微生物,在真菌中有木霉、漆斑霉、黑曲霉、青霉、根霉等;在细菌中有纤维粘菌属和纤维杆菌属;在放线菌中有黑红旋丝放线菌、玫瑰色放线菌、纤维放线菌及白玫瑰放线菌等。在已发现的产纤维素酶的菌株中,分解天然纤维素的能力较弱,酶活力不高,因此纤维素的利用受到一定的限制。自然界中存在有大量的纤维素资源可供人类所利用,运用现代科学技术发现和分离具有高活性的纤维素酶的微生物,对世界文明进步具有重要意义。

17
三、 果胶酶与果胶的降解 能够催化果胶酸(多聚半乳糖醛酸)或果胶(多聚甲氧基半乳糖醛酸)分子降解的酶类统称为果胶酶。果胶酶是当今开发生产的五大工业酶制剂之一,主要依靠真菌(霉菌)发酵生产。果胶酶普遍存在于植物和微生物中,人和动物不能合成果胶酶。果胶酶主要应用在工业生产中,以解除果胶质在果品加工过程及产品中所产生的不良影响。此外,果胶酶在果皮、果渣废料再生等方面也有应用。
或果胶(多聚甲氧基半乳糖醛酸)分子降解的酶类统称为果胶酶。果胶酶是当今开发生产的五大工业酶制剂之一,主要依靠真菌(霉菌)发酵生产。果胶酶普遍存在于植物和微生物中,人和动物不能合成果胶酶。果胶酶主要应用在工业生产中,以解除果胶质在果品加工过程及产品中所产生的不良影响。此外,果胶酶在果皮、果渣废料再生等方面也有应用。")
18
果胶酶根据其作用机理可以分为裂解酶和水解酶两类

19
四、糖原磷酸化酶与糖原的降解 糖原是葡萄糖残基以a-1,4-葡糖苷键连接带有分支的高聚体,糖原主要由肝脏和骨骼肌作为贮能而贮存。肌肉中贮存的糖原为延长肌肉收缩提供能量来源;肝脏中的糖原用于维持血糖水平。

20
糖原的降解是由糖原磷酸化酶(glycogen phosphorylase)和糖原脱支酶(glycogen-debranching enzyme)共同作用的。糖原磷酸化酶从糖原分子的非还原末端依次水解葡萄糖单元,作用键是a-1,4-葡糖苷键,产物为1-磷酸葡萄糖。由于该酶不能作用于a-1,6-葡糖苷键,所以不能完全降解糖原分子,残剩的具有分支部分称为糖原磷酸化酶界限糊精。
和糖原脱支酶(glycogen-debranching enzyme)共同作用的。糖原磷酸化酶从糖原分子的非还原末端依次水解葡萄糖单元,作用键是a-1,4-葡糖苷键,产物为1-磷酸葡萄糖。由于该酶不能作用于a-1,6-葡糖苷键,所以不能完全降解糖原分子,残剩的具有分支部分称为糖原磷酸化酶界限糊精。")
21
糖原脱支酶可作用于a-1,6-葡糖苷键,糖原磷酸化酶界限糊精经脱支酶水解,产物为葡萄糖和另一直链糊精,这一直链糊精又可被糖原磷酸化酶水解。糖原经磷酸化酶和脱支酶的协同作用,水解生成的1-磷酸葡萄糖和少量葡萄糖进入葡萄糖降解途径。

22
第二节 葡萄糖的降解 高等动物、植物和绝大多数微生物都能利用葡萄糖作碳源和能源,因此,葡萄糖的分解代谢,包括物质转化和能量转化规律,具有生物学的普遍意义。

23
碳水化合物进入酵解途径的前奏 除葡萄糖以外,其他碳水化合物通过酵解进入分解代谢,必须首先转变为酵解途径的任一中间物。 最重要的几种糖类化合物
1.贮存多糖(淀粉和糖原)、 2.二糖(麦芽糖、乳糖、蔗糖、海藻糖) 3.单糖(果糖、甘露糖、半乳糖)。
、 2.二糖(麦芽糖、乳糖、蔗糖、海藻糖) 3.单糖(果糖、甘露糖、半乳糖)。")
24
一、糖酵解(glycolysis)途径(EMP)
糖酵解是一系列反应,其反应过程发生在所有原核细胞和真核细胞的胞质溶胶中。糖酵解是指葡萄糖在一系列酶的催化下,经1,6-二磷酸果糖和3-磷酸甘油酸降解,生成丙酮酸并产生ATP的代谢过程。糖酵解是动物、植物、微生物细胞中普遍存在的葡萄糖降解途径,有氧或无氧条件下都能进行。一个葡萄糖分子经过酵解可转化为2分子丙酮酸。在糖酵解途径中,2个ATP分子需要提供给早期反应,糖酵解后期,有4个ATP生成。糖酵解有双重作用,首先是能量再生,其次是生成中间体,为许多生物合成途径提供前体。

25
糖酵解途径从葡萄糖到丙酮酸共由十步反应组成,分别由十种酶催化。这些酶全部在细胞液中,组成了可溶性的多酶体系。反应过程大体可分为三个阶段,第一阶段为葡萄糖经磷酸化生成1,6-二磷酸果糖,此阶段为糖的活化过程,主要变化是磷酸化及异构化;第二阶段是1,6-二磷酸果糖(二磷酸己糖)裂解为2分子磷酸丙糖,这是酵解过程中的一个关键步骤,葡萄糖的分解反应实际上是从这一步开始的,由于酵解是从双磷酸己糖开始降解的,故酵解作用又称双磷酸己糖途径(HDP途径);第三阶段是丙酮酸的生成,这一阶段的特点是高能磷酸键的生成(即ATP的生成)。
裂解为2分子磷酸丙糖,这是酵解过程中的一个关键步骤,葡萄糖的分解反应实际上是从这一步开始的,由于酵解是从双磷酸己糖开始降解的,故酵解作用又称双磷酸己糖途径(HDP途径);第三阶段是丙酮酸的生成,这一阶段的特点是高能磷酸键的生成(即ATP的生成)。")
26
(glucose-6-phosphate, G-6-P)
F-6-P F-1,6-2P ATP ADP 1,3-二磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸 丙酮酸 磷酸二 羟丙酮 3-磷酸 甘油醛 NAD+ NADH+H+ 磷酸烯醇式丙酮酸 (一)葡萄糖分解成丙酮酸 1、 葡萄糖磷酸化为6-磷酸葡萄糖 葡萄糖 6-磷酸葡萄糖 (glucose-6-phosphate, G-6-P) ATP ADP Mg2+ 己糖激酶 (hexokinase) (glucose)
葡萄糖分解成丙酮酸. 1、 葡萄糖磷酸化为6-磷酸葡萄糖. 葡萄糖. 6-磷酸葡萄糖. (glucose-6-phosphate, G-6-P) ATP ADP. Mg2+ 己糖激酶. (hexokinase) (glucose)")
27
反应要点: 第一步磷酸化反应不可逆;Go´= -16.7 kJ/ mol (ATP酸酐键:-30.54, 磷酸酯键:-13.81)(放能)
需要Mg 2+,ATP提供磷酸化基团 己糖激酶两个亚基构成,有4种同工酶,分Ⅰ、II、III和Ⅳ型, Ⅳ型存在肝细胞中,又称葡萄糖激酶(glucokinase,GK)。 GK的特点是: 对葡萄糖的亲和力很低(Km=10mmol/L,而HK的Km=0.1mmol/L) 受胰岛素的诱导 血糖调控中发挥重要作用 葡萄糖磷酸化的作用:极性增强不易透过脂膜失散;激活葡萄糖使之有利于与酶结合催化
。 GK的特点是: 对葡萄糖的亲和力很低(Km=10mmol/L,而HK的Km=0.1mmol/L) 受胰岛素的诱导. 血糖调控中发挥重要作用. 葡萄糖磷酸化的作用:极性增强不易透过脂膜失散;激活葡萄糖使之有利于与酶结合催化.")
28
醛糖和酮糖间的异构反应,碳1位形成自由羟基为下一步磷酸化准备 反应可逆,Go´=1.67kJ/mol
Glu G-6-P F-6-P F-1,6-2P ATP ADP 1,3-二磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸 丙酮酸 磷酸二 羟丙酮 3-磷酸 甘油醛 NAD+ NADH+H+ 磷酸烯醇式丙酮酸 2、6-磷酸葡萄糖转变为 6-磷酸果糖 6-磷酸葡萄糖 6-磷酸果糖 (fructose-6-phosphate, F-6-P) 磷酸己糖异构酶(phosphohexose isomerase) 醛糖和酮糖间的异构反应,碳1位形成自由羟基为下一步磷酸化准备 反应可逆,Go´=1.67kJ/mol
 磷酸己糖异构酶(phosphohexose isomerase) 醛糖和酮糖间的异构反应,碳1位形成自由羟基为下一步磷酸化准备. 反应可逆,Go´=1.67kJ/mol.")
29
1,6-双磷酸果糖 (1, 6-fructose-biphosphate, F-1,6-2P)
Glu G-6-P F-6-P F-1,6-2P ATP ADP 1,3-二磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸 丙酮酸 磷酸二 羟丙酮 3-磷酸 甘油醛 NAD+ NADH+H+ 磷酸烯醇式丙酮酸 3、 6-磷酸果糖转变为1,6-双磷酸果糖 1,6-双磷酸果糖 (1, 6-fructose-biphosphate, F-1,6-2P) 6-磷酸果糖 ATP ADP Mg2+ 6-磷酸果糖激酶-1 (6-phosphofructokinase-1)
 6-磷酸果糖. ATP ADP. Mg2+ 6-磷酸果糖激酶-1. (6-phosphofructokinase-1)")
30
反应特点 第二步磷酸化反应,不可逆反应;Go´= -14.23 kJ/mol(放能) 反应需要Mg 2+,ATP提供磷酸化基团
6-磷酸果糖激酶-1 变构酶,活性很低,是糖酵解途径中最重要的调节关键酶 别构激活剂:AMP; ADP; F-1,6-BP; F-2,6-BP 别构抑制剂: 柠檬酸, ATP, H+,2,3-二磷酸甘油酸 四个亚基组成,三种同工酶(A,B,C)
")
31
Dihydroxyacetone phosphate Glyceraldehyde -3-phosphate
Glu G-6-P F-6-P F-1,6-2P ATP ADP 1,3-二磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸 丙酮酸 磷酸二 羟丙酮 3-磷酸 甘油醛 NAD+ NADH+H+ 磷酸烯醇式丙酮酸 4、 磷酸己糖裂解成2分子磷酸丙糖 磷酸二羟丙酮 3-磷酸甘油醛 + 1,6-双磷酸果糖 Dihydroxyacetone phosphate 醛缩酶 (aldolase) Glyceraldehyde phosphate 吸能反应, Go´= kJ/mol,在标准条件下反应向缩合进行,但在细胞内却向裂解方向进行 醛缩酶属于第四类的裂合酶
 Glyceraldehyde -3-phosphate. 吸能反应, Go´= kJ/mol,在标准条件下反应向缩合进行,但在细胞内却向裂解方向进行. 醛缩酶属于第四类的裂合酶.")
32
5、磷酸丙糖的同分异构化 磷酸丙糖异构酶 (triose phosphate isomerase) 3-磷酸甘油醛 磷酸二羟丙酮
Glu G-6-P F-6-P F-1,6-2P ATP ADP 1,3-二磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸 丙酮酸 磷酸二 羟丙酮 3-磷酸 甘油醛 NAD+ NADH+H+ 磷酸烯醇式丙酮酸 5、磷酸丙糖的同分异构化 磷酸二羟丙酮 3-磷酸甘油醛 磷酸丙糖异构酶 (triose phosphate isomerase) Dihydroxyacetone phosphate Glyceraldehyde phosphate 磷酸丙糖异构酶催化反应是极其迅速的,反应平衡点偏向磷酸二羟丙酮,但由于3-磷酸甘油醛不断被消耗,前者不断转变为后者
 Dihydroxyacetone phosphate. Glyceraldehyde -3-phosphate. 磷酸丙糖异构酶催化反应是极其迅速的,反应平衡点偏向磷酸二羟丙酮,但由于3-磷酸甘油醛不断被消耗,前者不断转变为后者.")
33
(glyceraldehyde-3-phosphate dehydrogenase)
Glu G-6-P F-6-P F-1,6-2P ATP ADP 1,3-二磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸 丙酮酸 磷酸二 羟丙酮 3-磷酸 甘油醛 NAD+ NADH+H+ 磷酸烯醇式丙酮酸 6、3-磷酸甘油醛氧化为1,3-二磷酸甘油酸 1,3-二磷酸 甘油酸 3-磷酸甘油醛 Pi、NAD+ NADH+H+ 3-磷酸甘油醛脱氢酶 H (glyceraldehyde-3-phosphate dehydrogenase) 1,3-biphosphoglycerate 3-磷酸甘油醛脱氢酶催化醛基氧化为羧基和羧基磷酸化反应, 脱氢氧化,NAD+为受氢体,生成NADH+H+ 放能吸能偶联:放能 Go´= kJ/ mol;羧基磷酸化形成混合酸酐键是吸能 Go´= kJ/mol
 1,3-biphosphoglycerate. 3-磷酸甘油醛脱氢酶催化醛基氧化为羧基和羧基磷酸化反应, 脱氢氧化,NAD+为受氢体,生成NADH+H+ 放能吸能偶联:放能 Go´= kJ/ mol;羧基磷酸化形成混合酸酐键是吸能 Go´= kJ/mol.")
34
维生素PP NAD为受氢体的的氧化还原反应: AH2 + NAD+ A + NADH + H+

35
(phosphoglycerate kinase)
Glu G-6-P F-6-P F-1,6-2P ATP ADP 1,3-二磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸 丙酮酸 磷酸二 羟丙酮 3-磷酸 甘油醛 NAD+ NADH+H+ 磷酸烯醇式丙酮酸 7、1,3-二磷酸甘油酸转变成3-磷酸甘油酸 1,3-二磷酸 甘油酸 3-磷酸甘油酸 ADP Mg 2+ ATP 磷酸甘油酸激酶 (phosphoglycerate kinase) 第一次产生ATP,磷酸甘油酸激酶催化混合酸酐的磷酸基团转移到ADP,反应需Mg 2+ , 放能反应。 底物水平磷酸化:由底物分子反应释放能量或磷酸基团直接转移,使ADP( N DP)磷酸化生成ATP(NTP)的过程。
 第一次产生ATP,磷酸甘油酸激酶催化混合酸酐的磷酸基团转移到ADP,反应需Mg 2+ , 放能反应。 底物水平磷酸化:由底物分子反应释放能量或磷酸基团直接转移,使ADP( N DP)磷酸化生成ATP(NTP)的过程。")
36
(phosphoglycerate mutase)
Glu G-6-P F-6-P F-1,6-2P ATP ADP 1,3-二磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸 丙酮酸 磷酸二 羟丙酮 3-磷酸 甘油醛 NAD+ NADH+H+ 磷酸烯醇式丙酮酸 8、3-磷酸甘油酸转变为2-磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸 Mg 2+ 磷酸甘油酸 变位酶 (phosphoglycerate mutase)
")
37
烯醇磷酸酯具有高基团转移势能:Go´= -61.92 kJ/ mol
Glu G-6-P F-6-P F-1,6-2P ATP ADP 1,3-二磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸 丙酮酸 磷酸二 羟丙酮 3-磷酸 甘油醛 NAD+ NADH+H+ 磷酸烯醇式丙酮酸 9、2-磷酸甘油酸转变为磷酸烯醇式丙酮酸 + H2O 磷酸烯醇式丙酮酸 (phosphoenolpyruvate, PEP) 2-磷酸甘油酸 Mg 2+ 烯醇化酶 (enolase) 烯醇磷酸酯具有高基团转移势能:Go´= kJ/ mol
 2-磷酸甘油酸. Mg 2+ 烯醇化酶. (enolase) 烯醇磷酸酯具有高基团转移势能:Go´= kJ/ mol.")
38
高度放能反应,不可逆(Go´: -61.92对31.38 kJ/mol) 丙酮酸激酶又一个重要变构酶
Glu G-6-P F-6-P F-1,6-2P ATP ADP 1,3-二磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸 丙酮酸 磷酸二 羟丙酮 3-磷酸 甘油醛 NAD+ NADH+H+ 磷酸烯醇式丙酮酸 10、磷酸烯醇式丙酮酸转变成丙酮酸 磷酸烯醇式丙酮酸 丙酮酸 ADP ATP K+ Mg2+ 丙酮酸激酶 (pyruvate kinase) pyruvate phosphoenolpyruvate 第二次通过底物水平磷酸化生成ATP 高度放能反应,不可逆(Go´: 对31.38 kJ/mol) 丙酮酸激酶又一个重要变构酶
 pyruvate. phosphoenolpyruvate. 第二次通过底物水平磷酸化生成ATP. 高度放能反应,不可逆(Go´: 对31.38 kJ/mol) 丙酮酸激酶又一个重要变构酶.")
39
Lactate dehydrogenase
(二) 丙酮酸转变成乳酸 乳酸脱氢酶(LDH) NADH + H+ NAD+ Lactate dehydrogenase 丙酮酸 乳酸 缺氧条件下,丙酮酸还原为乳酸 反应中的NADH+H+ 来自于上述第6步反应中的 3-磷酸甘油醛脱氢反应。 LDH由四个亚基组成,五种同工每,每一种同工酶对底物有不同的亲和力
 丙酮酸转变成乳酸. 乳酸脱氢酶(LDH) NADH + H+ NAD+ Lactate dehydrogenase. 丙酮酸. 乳酸. 缺氧条件下,丙酮酸还原为乳酸. 反应中的NADH+H+ 来自于上述第6步反应中的 3-磷酸甘油醛脱氢反应。 LDH由四个亚基组成,五种同工每,每一种同工酶对底物有不同的亲和力.")
40
糖酵解的代谢途径 Glu G-6-P F-6-P F-1, 6-2P 1,3-二磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸 丙 酮 酸
E1 E2 Glu G-6-P F-6-P F-1, 6-2P ATP ADP 1,3-二磷酸甘油酸 3-磷酸甘油酸 2-磷酸甘油酸 丙 酮 酸 磷酸二羟丙酮 3-磷酸甘油醛 NAD+ NADH+H+ 磷酸烯醇式丙酮酸 糖酵解的代谢途径 E1:己糖激酶 E2: 6-磷酸果糖激酶-1 E3: 丙酮酸激酶 NADH+H+ NAD+ 乳 酸 E3

41
(三)糖酵解小结 反应部位:胞浆 糖酵解过程共进行11步反应,11个酶催化,有三步不可逆的反应(调控点) G G-6-P ATP ADP
己糖激酶 ATP ADP F-6-P F-1,6-2P 磷酸果糖激酶-1 ADP ATP PEP 丙酮酸 丙酮酸激酶

42
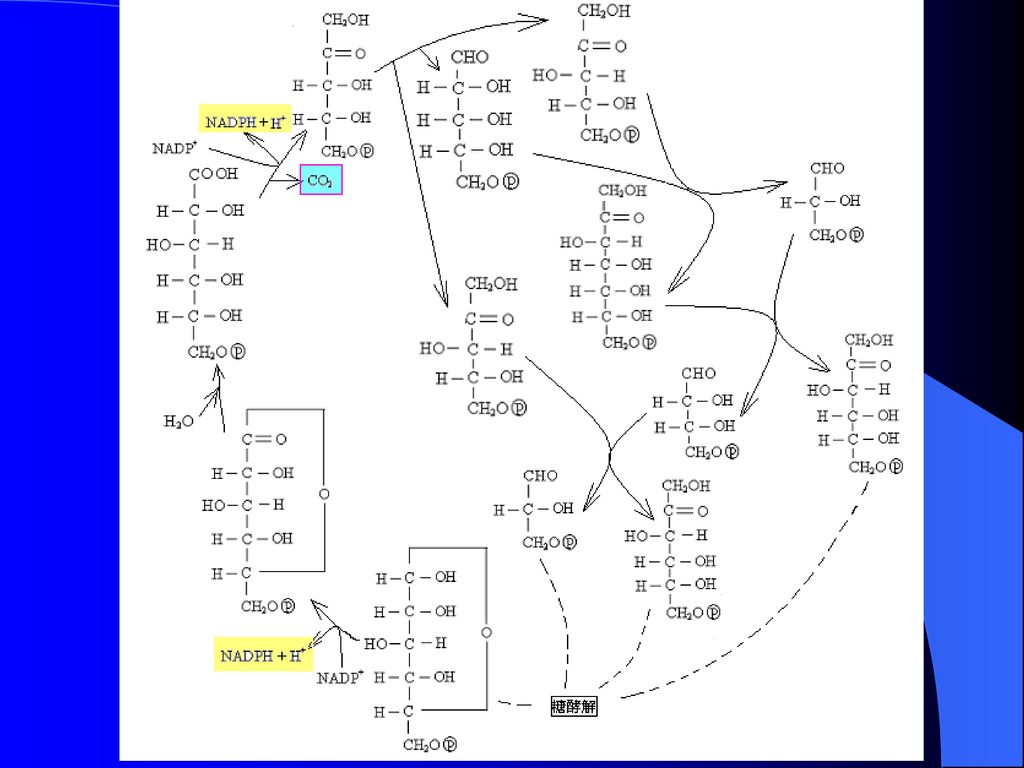
1分子葡萄糖分解为2分子乳酸的总反应和能量变化 C6H12O6 + 2ADP + 2Pi 2C3H6O3 + 2ATP +2H2O
糖酵解是一个不需氧的少量产能过程 产能方式:底物水平磷酸化 净生成ATP数量:从G开始 2×2-2= 2ATP 1分子葡萄糖分解为2分子乳酸的总反应和能量变化 C6H12O6 + 2ADP + 2Pi 2C3H6O3 + 2ATP +2H2O 终产物乳酸的去路 释放入血,进入肝脏再进一步代谢

43
(1) 葡萄糖被ATP磷酸化形成6-磷酸葡萄糖,催化此反应的酶是己糖激酶(hexokinase)
 葡萄糖被ATP磷酸化形成6-磷酸葡萄糖,催化此反应的酶是己糖激酶(hexokinase)")
44
(2) 6-磷酸葡萄糖被磷酸葡萄糖异构酶(phosphoglucoisomerase)转化为6-磷酸果糖,这种异构化反应包括醛糖转化为酮糖。
 6-磷酸葡萄糖被磷酸葡萄糖异构酶(phosphoglucoisomerase)转化为6-磷酸果糖,这种异构化反应包括醛糖转化为酮糖。")
45
(3) 6-磷酸果糖被ATP磷酸化生成1,6-二磷酸果糖,催化此反应的酶是磷酸果糖激酶(phosphofructokinase,PFK)。
 6-磷酸果糖被ATP磷酸化生成1,6-二磷酸果糖,催化此反应的酶是磷酸果糖激酶(phosphofructokinase,PFK)。")
46
(4) 1,6-二磷酸果糖被醛缩酶(Aldolase)作用裂解成2个三碳分子,分别是3-磷酸甘油醛和磷酸二羟丙酮。
 1,6-二磷酸果糖被醛缩酶(Aldolase)作用裂解成2个三碳分子,分别是3-磷酸甘油醛和磷酸二羟丙酮。")
47
(5) 丙糖磷酸异构酶(triose phosphate isomerase)催化3-磷酸甘油醛和磷酸二羟丙酮的相互转化。
 丙糖磷酸异构酶(triose phosphate isomerase)催化3-磷酸甘油醛和磷酸二羟丙酮的相互转化。")
48
(6) 3-磷酸甘油醛转化为1,3-二磷酸甘油酸。
 3-磷酸甘油醛转化为1,3-二磷酸甘油酸。")
49
(7) 1,3-二磷酸甘油酸新产生的高能磷酸键用于合成ATP,磷酸甘油酸激酶催化磷酰基从1,3-二磷酸甘油酸转移到ADP,再生ATP和3-磷酸甘油酸。
 1,3-二磷酸甘油酸新产生的高能磷酸键用于合成ATP,磷酸甘油酸激酶催化磷酰基从1,3-二磷酸甘油酸转移到ADP,再生ATP和3-磷酸甘油酸。")
50
(8) 3-磷酸甘油酸被磷酸甘油酸变位酶(phosphoglycerate mutase)转化为2-磷酸甘油酸。此反应是在同一分子内磷酸基到不同碳原子的移动。
 3-磷酸甘油酸被磷酸甘油酸变位酶(phosphoglycerate mutase)转化为2-磷酸甘油酸。此反应是在同一分子内磷酸基到不同碳原子的移动。")
51
(9)烯醇化酶(enolase)催化2-磷酸甘油酸的脱氢反应形成磷酸丙酮酸(PEP)。此反应将低能的2-磷酸甘油酸的磷酯键转化为PEP的高能磷酸键。
烯醇化酶(enolase)催化2-磷酸甘油酸的脱氢反应形成磷酸丙酮酸(PEP)。此反应将低能的2-磷酸甘油酸的磷酯键转化为PEP的高能磷酸键。")
52
(10)在最后反应中,丙酮酸激酶(pyruvate kinase)催化磷酰基从PEP到ADP形成ATP和丙酮酸,这是生理的不可逆转化过程。
在最后反应中,丙酮酸激酶(pyruvate kinase)催化磷酰基从PEP到ADP形成ATP和丙酮酸,这是生理的不可逆转化过程。")
53
从葡萄糖开始酵解生成丙酮酸的总反应式为:

55
糖酵解、柠檬酸循环和氧化磷酸化反应途径的亚细胞定位
lactate

56
所以,糖酵解是三羧酸循环和氧化磷酸化的前奏。如果供氧不足,NADH不进入呼吸链,而是把丙酮酸还原成乳酸。

57
二、丙酮酸(pyruvic acid)的无氧降解
在糖酵解过程中生成的丙酮酸和还原型辅酶都不是最终代谢产物,它们的去路因不同条件和不同生物而异。在有氧条件下和无氧条件下其代谢的最终产物也有所不同。本小节讨论无氧条件下丙酮酸的去路。

58
(1)同型乳酸(lactic acid)发酵
在糖酵解过程中,NAD+是使糖酵解继续进行的条件(1,3-二磷酸甘油酸的生成)。在有氧条件下,NAD+由NADH经电子传递链重新氧化而再生;当氧供给不足时,则NAD+的生成不足以维持糖酵解的继续进行,在这些条件下,丙酮酸作为受氢体,在乳酸脱氢酶(lactate dehydrogenase)的催化下,丙酮酸转化为乳酸(哺乳动物的骨骼肌激烈运动后产生的酸楚感即缘于此)。
。在有氧条件下,NAD+由NADH经电子传递链重新氧化而再生;当氧供给不足时,则NAD+的生成不足以维持糖酵解的继续进行,在这些条件下,丙酮酸作为受氢体,在乳酸脱氢酶(lactate dehydrogenase)的催化下,丙酮酸转化为乳酸(哺乳动物的骨骼肌激烈运动后产生的酸楚感即缘于此)。")
59
同型乳酸发酵就是利用乳酸菌能产生活性很强的乳酸脱氢酶,在无氧条件下,将丙酮酸还原成乳酸。
由葡萄糖进行乳酸发酵的总反应式如下(未计ADP磷酸化的水分子出入):
:")
60
由反应式可知,1分子葡萄糖经乳酸发酵可生成2分子乳酸,共释放化学能196.5kJ/mol,同时生成2个ATP,能量利用率为34.0%。
同型发酵的乳酸菌类群主要有双球菌Diplococcus)、链球菌(Streplococcus)、乳杆菌(Lactobacillus)等兼性微生物。L-型乳酸是重要的化工原料和食品酸味剂,工业上一般采用以淀粉质原料进行发酵生产。 反应中的NADH+H+ 来自于上述第6步反应中的 3-磷酸甘油醛脱氢反应。
、链球菌(Streplococcus)、乳杆菌(Lactobacillus)等兼性微生物。L-型乳酸是重要的化工原料和食品酸味剂,工业上一般采用以淀粉质原料进行发酵生产。 反应中的NADH+H+ 来自于上述第6步反应中的 3-磷酸甘油醛脱氢反应。")
61
(2)乙醇(alcohol)发酵 在酵母和一些其它微生物中,无氧条件下为使糖酵解继续进行所要求的NAD+由乙醇发酵过程再生。酵母细胞不仅能产生糖酵解途径的全部酶系,还能产生丙酮酸脱羧酶(pyruvate decarboxylase)和乙醇脱氢酶(alcohol dehydrogenase)。酵母菌在无氧条件下将葡萄糖酵解产生丙酮酸,丙酮酸脱羧酶以焦磷酸硫胺素(TPP)作为辅酶,催化丙酮酸脱羧,生成乙醛;乙醇脱氢酶以NADH为辅酶,将乙醛还原生成乙醇。
和乙醇脱氢酶(alcohol dehydrogenase)。酵母菌在无氧条件下将葡萄糖酵解产生丙酮酸,丙酮酸脱羧酶以焦磷酸硫胺素(TPP)作为辅酶,催化丙酮酸脱羧,生成乙醛;乙醇脱氢酶以NADH为辅酶,将乙醛还原生成乙醇。")
63
由葡萄糖进行乙醇发酵的总反应式如下(未计ADP磷酸化的水分子出入):
酒精发酵是酵母菌在无氧条件下分解葡萄糖取得生物能量的代谢方式,释放的化学能为234.3kj/mol,净生成2个ATP,能量利用率为28.6%。

64
(3)甘油(glycerin)发酵 正常的乙醇发酵会产生少量甘油,这是因为乙醇发酵之初,细胞内没有足够的乙醛作为受氢体,致使NADH浓度升高,被a-磷酸甘油脱氢酶用于磷酸二羟丙酮的还原反应,生成a-磷酸甘油,NADH被氧化成NAD+。a-磷酸甘油则在磷酯酶的水解作用下生成甘油。发酵过程中将受氢体乙醛去除作为控制发酵的条件,从而使发酵液中积累甘油,这是酵母菌甘油发酵的基本原理。酵母菌甘油发酵有下列两种方法: 亚硫酸盐法甘油发酵 和 碱法甘油发酵

65
亚硫酸盐法甘油发酵 酵母酒精发酵时,在发酵液中加入亚硫酸氢钠,乙醛与亚硫酸氢钠起加成反应,生成难溶的结晶状乙醛亚硫酸钠加成物。

66
乙醛的去除为磷酸二羟丙酮的还原创造条件。

67
a-磷酸甘油在 a-磷酸甘油磷酸酯酶的作用下水解生成甘油。

68
自葡萄糖开始的总反应式为: 从式中可以看出,1分子葡萄糖理论上只可以生成1分子甘油,反应中无ATP生成,故在反应中应控制亚硫酸的量,适当保留部分乙醇发酵,以维持酵母生长、发酵所需的能量。

69
碱法甘油发酵 酵母酒精发酵时,使发酵液保持碱性(pH7.6),乙醛之间起歧化反应,相互氧化还原,生成等量的乙醇和乙酸。
,乙醛之间起歧化反应,相互氧化还原,生成等量的乙醇和乙酸。")
70
乙醛失去了作为受氢体的作用,NADH只能用于磷酸二羟丙酮的还原,生成甘油。自葡萄糖开始的总反应式为:
反应式表明,碱法甘油发酵同样不能产生ATP为细胞提供能量,只能用大量酵母在非生长情况下进行甘油发酵。

71
三、糖酵解能量收率 无氧

72
有氧

73
四、其他单糖的降解 果糖的降解 果糖是人类广泛食用的糖,果糖也是大多数异养微生物能吸收利用的一种己糖,一般在自然界中分布广泛,以游离状态存在于植物中。果糖的降解通常有两种情况:

74
(1)果糖经6-磷酸果糖的降解 (2)果糖经1-磷酸果糖的降解
果糖经6-磷酸果糖的降解 (2)果糖经1-磷酸果糖的降解")
75
半乳糖的降解 1)半乳糖被半乳糖激酶(galactokinase)磷酸化得到1-磷酸半乳糖。 2)1-磷酸半乳糖尿苷转移酶(galactose 1-phosphate uridylyl transferase)催化尿苷基从UDP-葡萄糖到1-磷酸半乳糖的转移形成UDP-半乳糖和1-磷酸葡萄糖。 3)UDP-半乳糖被UDP-半乳糖4-差向异构酶(UDP-galactose 4-epimerase)转回到UDP-葡萄糖,因而,UDP-葡萄糖在反应途径中并未消耗。 4)最后,1-磷酸葡萄糖被磷酸葡萄糖变位酶(phosphoglucomutase)转化为6-磷酸葡萄糖,进入EMP途径。
UDP-半乳糖被UDP-半乳糖4-差向异构酶(UDP-galactose 4-epimerase)转回到UDP-葡萄糖,因而,UDP-葡萄糖在反应途径中并未消耗。 4)最后,1-磷酸葡萄糖被磷酸葡萄糖变位酶(phosphoglucomutase)转化为6-磷酸葡萄糖,进入EMP途径。")
76
其他己糖的代谢 除葡萄糖外,其它己糖也可转变成磷酸己糖而进入酵解途径。 Glu G-6-P F-6-P F-1,6-2P 丙酮酸 半乳糖
1-磷酸半乳糖 1-磷酸葡萄糖 半乳糖激酶 变位酶 Glu G-6-P F-6-P F-1,6-2P ATP ADP 丙酮酸 甘露糖 6-磷酸甘露糖 己糖激酶 异构酶 鸟苷酰转移酶 4位差向异构酶 果糖 己糖激酶

77
五、糖酵解的调节 在糖酵解过程中有三个不可逆反应,也就是有三个调控步骤,分别由三种不同的酶调节,它们是磷酸果糖激酶、己糖激酶和丙酮酸激酶。

78
① 己糖激酶 ② 磷酸果糖激酶 ③ 丙酮酸激酶 关键酶 ① 变构调节 ② 共价修饰调节 调节方式

79
磷酸果糖激酶(phosphofructokinase, PFK) 磷酸果糖激酶是一个变构酶,其分子由四个亚基组成,该酶是酵解过程最关键的限速酶,由磷酸果糖激酶催化的反应步骤被称为糖酵解的限速步骤(committed step)。该酶的调节有多种途径: (1)ATP/AMP调节 ATP是磷酸果糖激酶的底物,也是变构调节物。高浓度的ATP会降低磷酸果糖激酶对6-磷酸果糖的亲和力。PFK被ATP变构抑制,但这种抑制作用被AMP逆转。当ATP供应短缺时(即AMP充足),反应速度加快,生成更多的ATP;当有足够多的ATP时,反应速度减慢。这一点具有重要的生理意义。 (2)柠檬酸调节 柠檬酸可增加ATP对PFK的抑制作用。高水平的柠檬酸是生物合成前体的碳骨架过剩的信号,葡萄糖就不进一步酵解。
ATP/AMP调节 ATP是磷酸果糖激酶的底物,也是变构调节物。高浓度的ATP会降低磷酸果糖激酶对6-磷酸果糖的亲和力。PFK被ATP变构抑制,但这种抑制作用被AMP逆转。当ATP供应短缺时(即AMP充足),反应速度加快,生成更多的ATP;当有足够多的ATP时,反应速度减慢。这一点具有重要的生理意义。 (2)柠檬酸调节 柠檬酸可增加ATP对PFK的抑制作用。高水平的柠檬酸是生物合成前体的碳骨架过剩的信号,葡萄糖就不进一步酵解。")
80
己糖激酶(hexokinase) 已糖激酶也是一个变构酶,该酶催化糖酵解的第一步不可逆反应,受到6-磷酸葡萄糖的抑制。当PFK被抑制时,其底物6-磷酸果糖浓度升高,而与6-磷酸果糖处于动态平衡的6-磷酸葡萄糖浓度相应升高,高浓度的6-磷酸葡萄糖对己糖激酶的活性实施抑制。因此,磷酸果糖激酶活性的抑制与己糖激酶活性的抑制是相互联系的,并且磷酸果糖激酶是主要的调控步骤。

81
丙酮酸激酶(pyruvate kinase)
在糖酵解中丙酮酸激酶催化第三个不可逆步骤,该酶也是一个变构酶,它被1,6-二磷酸果糖活化。ATP和丙氨酸变构地抑制此酶,当ATP的供给和生物合成前体(丙氨酸的水平)足够高时,磷酸烯醇式丙酮酸转变为丙酮酸的速度大大下降,糖酵解速度减慢。
足够高时,磷酸烯醇式丙酮酸转变为丙酮酸的速度大大下降,糖酵解速度减慢。")
82
六、糖酵解的生理意义 (1)糖酵解途径是单糖分解代谢的一条最重要的基本途径,该途径在各类生物中的分布最为广泛,并且在有氧或无氧条件下都能运转。其它糖类如己糖(果糖、半乳糖、甘露糖等)、戊糖都能通过特定的方式进入糖酵解途径。 (2)细胞在无氧条件下,通过糖酵解可以获得有限的能量用以维持生命。 (3)糖酵解过程为有氧条件下单糖的彻底氧化提供了充分的条件。同时为体内其它物质的合成提供原料。
细胞在无氧条件下,通过糖酵解可以获得有限的能量用以维持生命。 (3)糖酵解过程为有氧条件下单糖的彻底氧化提供了充分的条件。同时为体内其它物质的合成提供原料。")
83
第三节 丙酮酸的有氧降解 在有氧的条件下,大多数动植物及微生物都可将糖酵解产生的丙酮酸进行有氧降解,并释放更多的能量。在这一过程中,丙酮酸被丙酮酸脱氢酶系(pyruvate dehydrogenase)作用进行氧化脱羧反应(oxidative decarboxylation),丙酮酸经氧化脱羧形成乙酰-CoA 然后,乙酰-CoA与草酰乙酸结合进入三羧酸循环彻底氧化为水和二氧化碳,同时产生大量的ATP。
作用进行氧化脱羧反应(oxidative decarboxylation),丙酮酸经氧化脱羧形成乙酰-CoA. 然后,乙酰-CoA与草酰乙酸结合进入三羧酸循环彻底氧化为水和二氧化碳,同时产生大量的ATP。")
84
丙酮酸的有氧降解(即葡萄糖的有氧氧化)是指在有氧条件下,丙酮酸通过丙酮酸氧化脱羧酶系的作用生成乙酰CoA(acetyl coenzyme),并进入三羧酸循环(柠檬酸循环)彻底氧化为水和二氧化碳的过程。也就是说丙酮酸氧化脱羧生成乙酰-CoA的反应不是三羧酸循环中的反应,而是糖酵解和三羧酸循环之间的桥梁,真正进入三羧酸循环的是乙酰-CoA。这一复杂的过程分两段完成: 第一阶段:丙酮酸的氧化脱羧(丙酮酸——乙酰CoA)。 第二阶段:三羧酸循环(乙酰CoA与草酰乙酸结合进入TCA循环)。
。 第二阶段:三羧酸循环(乙酰CoA与草酰乙酸结合进入TCA循环)。")
85
有氧氧化的反应过程 G(Gn) 胞液 第一阶段:酵解途径 丙酮酸 第二阶段:丙酮酸的氧化脱羧 乙酰CoA 第三阶段:三羧酸循环 线粒体
第四阶段:氧化磷酸化 TAC循环 NADH+H+ FADH2 [O] CO2 H2O ADP ATP

86
一、丙酮酸氧化脱羧反应 丙酮酸氧化脱氢酶系是由丙酮酸脱氢酶(E1)、二氢硫辛酸转乙酰基酶(E2)、二氢硫辛酸脱氢酶(E3)这三种酶和TPP(焦磷酸硫胺素)、CoASH、硫辛酸、FAD、NAD+、Mg2+六种辅助因子组成的。丙酮酸氧化脱氢酶系是在线粒体膜上,催化反应如下:
、二氢硫辛酸转乙酰基酶(E2)、二氢硫辛酸脱氢酶(E3)这三种酶和TPP(焦磷酸硫胺素)、CoASH、硫辛酸、FAD、NAD+、Mg2+六种辅助因子组成的。丙酮酸氧化脱氢酶系是在线粒体膜上,催化反应如下:")
87
丙酮酸脱氢酶复合体的组成 酶 辅助因子 E1:丙酮酸脱氢酶 TPP E2:二氢硫辛酰胺转乙酰酶 硫辛酸 HSCoA
NAD+ 酶 辅助因子 E1:丙酮酸脱氢酶 TPP E2:二氢硫辛酰胺转乙酰酶 硫辛酸 HSCoA E3:二氢硫辛酰胺脱氢酶 FAD, NAD+

89
反应特点 反应部位:线粒体 反应性质 :氧化脱羧,产物为NADH+H+ 、CO2 和CH3CO~SCoA;不可逆反应(Go´=-39.5kJ/mol ) 丙酮酸脱氢酶复合体:三个酶和五个辅助因子(TPP、LA、HS~CoA、FAD、NAD+),连续催化,没有游离的中间产物,无副反应,该复合体活性受到调节。
,连续催化,没有游离的中间产物,无副反应,该复合体活性受到调节。")
90
二、丙酮酸的有氧降解——三羧酸循环(tricarboxylic acid, TCA, Krebs) (一)三羧酸循环途径
 (一)三羧酸循环途径")
92
TCA小结 TAC部位:线粒体。 三羧酸循环的要点: 消耗一分子乙酰CoA, 经四次脱氢,二次脱羧,一次底物水平磷酸化。
生成1分子FADH2,3分子NADH+H+,2分子CO2, 1分子GTP。 关键酶:柠檬酸合酶、α-酮戊二酸脱氢酶复合体、异柠檬酸脱氢酶 整个循环反应为不可逆反应

93
(二)三羧酸循环的调节 三羧酸循环存在着自动调节机制,其循环中的每一步反应都受到严密的控制。从乙酰CoA开始进入三羧酸循环,共存在着三个不可逆反应,它们分别是由柠檬酸合成酶、异柠檬酸脱氢酶和a-酮戊二酸脱氢酶系催化的反应,研究认为调控机制如下:
三羧酸循环的调节 三羧酸循环存在着自动调节机制,其循环中的每一步反应都受到严密的控制。从乙酰CoA开始进入三羧酸循环,共存在着三个不可逆反应,它们分别是由柠檬酸合成酶、异柠檬酸脱氢酶和a-酮戊二酸脱氢酶系催化的反应,研究认为调控机制如下:")
94
(1)柠檬酸合成酶是一个变构酶,其催化的反应是三羧酸循环中的第一步,可调节总反应速度,是关键反应步骤,因此该酶又称为限速酶。在高能荷的情况下,ATP降低柠檬酸合成酶对乙酰CoA的亲和力,酶活力下降;此外,NADH、脂酰CoA、琥珀酰CoA都对该酶有抑制作用,反应产物柠檬酸对该酶有反馈抑制作用。草酰乙酸的有效浓度同样保证三羧酸循环的正常进行。
柠檬酸合成酶是一个变构酶,其催化的反应是三羧酸循环中的第一步,可调节总反应速度,是关键反应步骤,因此该酶又称为限速酶。在高能荷的情况下,ATP降低柠檬酸合成酶对乙酰CoA的亲和力,酶活力下降;此外,NADH、脂酰CoA、琥珀酰CoA都对该酶有抑制作用,反应产物柠檬酸对该酶有反馈抑制作用。草酰乙酸的有效浓度同样保证三羧酸循环的正常进行。")
95
(2)异柠檬酸脱氢酶受到Ca2+和ADP的别构激活,ADP能增强异柠檬酸脱氢酶与异柠檬酸之间的亲和力。而ATP、NADH则对该酶的活性有抑制作用。
(3)a-酮戊二酸脱氢酶系中的二氢硫辛酸琥珀酰转移酶是使三羧酸进入二羧酸的关键酶,也是其它代谢产物进入三羧酸循环的关键,它能调节三羧酸循环的正常运行,并限制外来的a-酮戊二酸进入三羧酸循环。该酶的活性受其产物琥珀酰CoA的反馈抑制,NADH对该酶的活性有抑制作用。
a-酮戊二酸脱氢酶系中的二氢硫辛酸琥珀酰转移酶是使三羧酸进入二羧酸的关键酶,也是其它代谢产物进入三羧酸循环的关键,它能调节三羧酸循环的正常运行,并限制外来的a-酮戊二酸进入三羧酸循环。该酶的活性受其产物琥珀酰CoA的反馈抑制,NADH对该酶的活性有抑制作用。")
96
(三)三羧酸循环的能量生成 由三羧酸循环产生的NADH和FADH2被再氧化,通过氧化磷酸化释放出能量合成ATP,三羧酸循环每循环一次产生3个NADH分子,每个NADH分子通过氧化磷酸化途径生成2.5个ATP分子。而一分子FADH2则生成1.5个ATP分子。当琥珀酰CoA转变为琥珀酸时,直接合成一个GTP(或ATP)。因此一分子乙酰CoA通过三羧酸循环和氧化磷酸化共产生10个ATP分子。
。因此一分子乙酰CoA通过三羧酸循环和氧化磷酸化共产生10个ATP分子。")
98
(四)三羧酸循环的生理意义 (1)为细胞提供能量 糖在有氧条件下彻底氧化释放出的总能量远远大于糖的无氧酵解,所形成的ATP也多于糖的无氧酵解。循环代谢所产生的能量用于机体代谢的各个方面,诸如生物合成、物质主动运输、运动、生物发光等。

99
(2)三羧酸循环是机体代谢的枢纽 三羧酸循环即柠檬酸循环是绝大多数生物体主要的分解代谢途径,同时也是提供大量自由能的重要代谢系统;三羧酸循环具有分解代谢和合成代谢双重性或称两重性。三羧酸循环是联系各类物质代谢的枢纽。
三羧酸循环是机体代谢的枢纽 三羧酸循环即柠檬酸循环是绝大多数生物体主要的分解代谢途径,同时也是提供大量自由能的重要代谢系统;三羧酸循环具有分解代谢和合成代谢双重性或称两重性。三羧酸循环是联系各类物质代谢的枢纽。")
100
(3)三羧酸循环的产物可以得到最经济的利用 例如能量的利用、中间产物的利用等。循环中产生的CO2部分被用于某些物质的生物合成。
三羧酸循环的产物可以得到最经济的利用 例如能量的利用、中间产物的利用等。循环中产生的CO2部分被用于某些物质的生物合成。")
101
三、柠檬酸发酵 在生产过程中,利用微生物在特定条件下的固有代谢规律,自然积累某种产品的发酵方法称为自然发酵法。例如乙醇、乳酸等产品的生产,许多自然发酵产品都是微生物自身不能再利用的代谢产物,容易积累。根据微生物代谢途径及代谢的调节机制采取具有针对性的有效措施,改变微生物固有的代谢平衡,从而达到某一中间产物的积累,以提高产品得率,这种建立在代谢调控理论基础上的发酵技术称为代谢调节发酵。柠檬酸发酵即属此类。

103
柠檬酸是三羧酸循环的中间产物,若想利用EMP-TCA循环途径大量积累柠檬酸,应设法阻断柠檬酸的进一步反应。方法之一是针对催化柠檬酸进一步反应的顺乌头酸酶的酶学性质使用抑制剂。顺乌头酸酶是个含铁的非血红素蛋白,由铁硫中心(Fe4S4)作为辅基,催化底物脱水、加水反应。因此,在发酵过程中当菌体生长繁殖到足够数量时,适量加入亚铁氰化钾(黄血盐)与铁硫中心的Fe++络合生成络合物,则该酶活性丧失或活力大大降低,以达到柠檬酸积累的目的。方法之二是通过诱变造成生产菌种的顺乌头酸酶缺损或活力降低,同样可以积累柠檬酸。
作为辅基,催化底物脱水、加水反应。因此,在发酵过程中当菌体生长繁殖到足够数量时,适量加入亚铁氰化钾(黄血盐)与铁硫中心的Fe++络合生成络合物,则该酶活性丧失或活力大大降低,以达到柠檬酸积累的目的。方法之二是通过诱变造成生产菌种的顺乌头酸酶缺损或活力降低,同样可以积累柠檬酸。")
104
草酰乙酸是合成柠檬酸的前体,由于顺乌头酸催化的反应被阻断,三羧酸循环不能正常进行,也就无法保证草酰乙酸的生成,导致无法合成柠檬酸,因此必须解决草酰乙酸的来源。丙酮酸羧化支路是草酰乙酸的另一来源,应选用回补支路旺盛的菌种解决草酰乙酸的来源。 柠檬酸发酵所用的主要微生物为黑曲霉(Asp.niger),在黑曲霉中具有三羧酸循环和乙醛酸循环的酶系,并且黑曲霉能分泌淀粉酶,所以可以利用淀粉质原料直接生产柠檬酸。
,在黑曲霉中具有三羧酸循环和乙醛酸循环的酶系,并且黑曲霉能分泌淀粉酶,所以可以利用淀粉质原料直接生产柠檬酸。")
105
四、回补途径 三羧酸循环的中间产物可以作为某些生物合成的前体,而中间产物的被利用,势必导致循环的平衡受到影响,只有不断补充这些中间产物才能维持三羧酸循环的正常运行。这种补充中间产物的酶反应称为回补反应(anaplerotic reaction),正常情况下,三羧酸循环中间产物的动用、消耗和回补处于动态平衡中,其浓度几乎保持恒定。
,正常情况下,三羧酸循环中间产物的动用、消耗和回补处于动态平衡中,其浓度几乎保持恒定。")
106
(一)丙酮酸羧化支路 三碳酸固定CO2生成四碳酸的过程。
丙酮酸羧化支路 三碳酸固定CO2生成四碳酸的过程。")
108
(二)乙醛酸循环(glyoxylate cycle)
乙醛酸循环(glyoxylate cycle)又称乙醛酸途径(glyoxylate pathway),是指乙酰CoA的另一种去向,它借助于TCA循环的某些反应,最终生成乙醛酸。 乙醛酸循环从草酰乙酸与乙酰CoA缩合开始,形成柠檬酸后,异构化成异柠檬酸,异柠檬酸被异柠檬酸裂解酶(isocitrate lyase)裂解成琥珀酸和乙醛酸(乙醛酸循环因循环中的中间产物为乙醛酸而得名),乙醛酸与另一个乙酰CoA在苹果酸合成酶(malate synthetase)的催化下缩合成苹果酸,苹果酸氧化成草酰乙酸进而循环。
又称乙醛酸途径(glyoxylate pathway),是指乙酰CoA的另一种去向,它借助于TCA循环的某些反应,最终生成乙醛酸。 乙醛酸循环从草酰乙酸与乙酰CoA缩合开始,形成柠檬酸后,异构化成异柠檬酸,异柠檬酸被异柠檬酸裂解酶(isocitrate lyase)裂解成琥珀酸和乙醛酸(乙醛酸循环因循环中的中间产物为乙醛酸而得名),乙醛酸与另一个乙酰CoA在苹果酸合成酶(malate synthetase)的催化下缩合成苹果酸,苹果酸氧化成草酰乙酸进而循环。")
110
第四节 戊糖磷酸途径 一、戊糖磷酸途径 葡萄糖的有氧代谢EMP-TCA循环是各种生物体普遍存在的氧化分解途径,是主要的产能途径。然而研究发现,当用碘乙酸或氟化钠抑制酵解途径时,呼吸作用仍能消耗葡萄糖;用同位素14C分别标记葡萄糖C1和C6,如果糖酵解是唯一代谢途径,那么14C1和14C6生成14CO2的速度应该相同,但是,实验结果表明,14C1更容易氧化成14CO2。证明葡萄糖还有其它代谢途径。

111
戊糖磷酸途径(hexose monophosphate shunt, HMS[P]) (也称为己糖单磷酸旁路)或称磷酸葡萄糖酸途径(phosphogluconate pathway)就是动物、植物、微生物细胞中普遍存在的另一条重要的葡萄糖分解途径,这一途径代表了与还原型辅酶NADPH生成相偶联的葡萄糖氧化作用的另一种模型。戊糖磷酸途径是大多数细胞中生物合成反应的还原力的重要来源,其酶系存在于细胞液中,胞液是许多需要NADPH的生物合成反应的场所。该途径在己糖和戊糖的互变中是相当重要的,在戊糖的合成、分解代谢以及为光合作用中碳固定所需要的磷酸化戊糖核酮糖-1,5-二磷酸的再生中,这条途径起到了关键性作用。
![戊糖磷酸途径(hexose monophosphate shunt, HMS[P]) (也称为己糖单磷酸旁路)或称磷酸葡萄糖酸途径(phosphogluconate pathway)就是动物、植物、微生物细胞中普遍存在的另一条重要的葡萄糖分解途径,这一途径代表了与还原型辅酶NADPH生成相偶联的葡萄糖氧化作用的另一种模型。戊糖磷酸途径是大多数细胞中生物合成反应的还原力的重要来源,其酶系存在于细胞液中,胞液是许多需要NADPH的生物合成反应的场所。该途径在己糖和戊糖的互变中是相当重要的,在戊糖的合成、分解代谢以及为光合作用中碳固定所需要的磷酸化戊糖核酮糖-1,5-二磷酸的再生中,这条途径起到了关键性作用。](http://slidesplayer.com/slide/11369248/61/images/111/%E6%88%8A%E7%B3%96%E7%A3%B7%E9%85%B8%E9%80%94%E5%BE%84%EF%BC%88hexose+monophosphate+shunt%2C+HMS%5BP%5D%EF%BC%89+%EF%BC%88%E4%B9%9F%E7%A7%B0%E4%B8%BA%E5%B7%B1%E7%B3%96%E5%8D%95%E7%A3%B7%E9%85%B8%E6%97%81%E8%B7%AF%EF%BC%89%E6%88%96%E7%A7%B0%E7%A3%B7%E9%85%B8%E8%91%A1%E8%90%84%E7%B3%96%E9%85%B8%E9%80%94%E5%BE%84%EF%BC%88phosphogluconate+pathway%EF%BC%89%E5%B0%B1%E6%98%AF%E5%8A%A8%E7%89%A9%E3%80%81%E6%A4%8D%E7%89%A9%E3%80%81%E5%BE%AE%E7%94%9F%E7%89%A9%E7%BB%86%E8%83%9E%E4%B8%AD%E6%99%AE%E9%81%8D%E5%AD%98%E5%9C%A8%E7%9A%84%E5%8F%A6%E4%B8%80%E6%9D%A1%E9%87%8D%E8%A6%81%E7%9A%84%E8%91%A1%E8%90%84%E7%B3%96%E5%88%86%E8%A7%A3%E9%80%94%E5%BE%84%EF%BC%8C%E8%BF%99%E4%B8%80%E9%80%94%E5%BE%84%E4%BB%A3%E8%A1%A8%E4%BA%86%E4%B8%8E%E8%BF%98%E5%8E%9F%E5%9E%8B%E8%BE%85%E9%85%B6NADPH%E7%94%9F%E6%88%90%E7%9B%B8%E5%81%B6%E8%81%94%E7%9A%84%E8%91%A1%E8%90%84%E7%B3%96%E6%B0%A7%E5%8C%96%E4%BD%9C%E7%94%A8%E7%9A%84%E5%8F%A6%E4%B8%80%E7%A7%8D%E6%A8%A1%E5%9E%8B%E3%80%82%E6%88%8A%E7%B3%96%E7%A3%B7%E9%85%B8%E9%80%94%E5%BE%84%E6%98%AF%E5%A4%A7%E5%A4%9A%E6%95%B0%E7%BB%86%E8%83%9E%E4%B8%AD%E7%94%9F%E7%89%A9%E5%90%88%E6%88%90%E5%8F%8D%E5%BA%94%E7%9A%84%E8%BF%98%E5%8E%9F%E5%8A%9B%E7%9A%84%E9%87%8D%E8%A6%81%E6%9D%A5%E6%BA%90%EF%BC%8C%E5%85%B6%E9%85%B6%E7%B3%BB%E5%AD%98%E5%9C%A8%E4%BA%8E%E7%BB%86%E8%83%9E%E6%B6%B2%E4%B8%AD%EF%BC%8C%E8%83%9E%E6%B6%B2%E6%98%AF%E8%AE%B8%E5%A4%9A%E9%9C%80%E8%A6%81NADPH%E7%9A%84%E7%94%9F%E7%89%A9%E5%90%88%E6%88%90%E5%8F%8D%E5%BA%94%E7%9A%84%E5%9C%BA%E6%89%80%E3%80%82%E8%AF%A5%E9%80%94%E5%BE%84%E5%9C%A8%E5%B7%B1%E7%B3%96%E5%92%8C%E6%88%8A%E7%B3%96%E7%9A%84%E4%BA%92%E5%8F%98%E4%B8%AD%E6%98%AF%E7%9B%B8%E5%BD%93%E9%87%8D%E8%A6%81%E7.jpg "戊糖磷酸途径(hexose monophosphate shunt, HMS[P]) (也称为己糖单磷酸旁路)或称磷酸葡萄糖酸途径(phosphogluconate pathway)就是动物、植物、微生物细胞中普遍存在的另一条重要的葡萄糖分解途径,这一途径代表了与还原型辅酶NADPH生成相偶联的葡萄糖氧化作用的另一种模型。戊糖磷酸途径是大多数细胞中生物合成反应的还原力的重要来源,其酶系存在于细胞液中,胞液是许多需要NADPH的生物合成反应的场所。该途径在己糖和戊糖的互变中是相当重要的,在戊糖的合成、分解代谢以及为光合作用中碳固定所需要的磷酸化戊糖核酮糖-1,5-二磷酸的再生中,这条途径起到了关键性作用。")
113
HMS途径的生理意义 (1)产生NADPH为生物合成提供还原力 (2)产生戊糖磷酸参加核酸代谢 (3)NADPH使红细胞中还原型谷胱甘肽再生,对维持红细胞还原性有重要作用。 (4)戊糖磷酸途径是植物光合作用中从CO2合成葡萄糖的部分途径,同时也是戊糖代谢的主要途径。其代谢方式一般是以戊糖磷酸形式进入途径,进而与EMP、TCA循环等代谢途径相联结。 (5)在特殊情况下,戊糖磷酸途径也可为细胞提供能量
戊糖磷酸途径是植物光合作用中从CO2合成葡萄糖的部分途径,同时也是戊糖代谢的主要途径。其代谢方式一般是以戊糖磷酸形式进入途径,进而与EMP、TCA循环等代谢途径相联结。 (5)在特殊情况下,戊糖磷酸途径也可为细胞提供能量.")
114
HMS途径的能量生成 NADPH的电子转交给NAD+,经呼吸链氧化产生能量,按氧化1分子葡萄糖计算,可产生30分子ATP,扣除开始消耗的1分子ATP,净得29分子ATP。产能与EMP-TCA循环途径相当。

115
二、磷酸解酮酶途径(PK)与异型乳酸发酵
与异型乳酸发酵")
116
异型乳酸发酵是糖经PK途径进行的发酵作用,产物除乳酸外,还有比例较高的乙醇和二氧化碳。进行异型乳酸发酵的微生物有肠膜状明串珠菌(Leuconostoc mesenteroides)、番茄乳杆菌(Lac. lycopersici)、短乳杆菌(Lactobacillus brevis)、甘露醇乳杆菌(Lac. manitopoeum)、双歧杆菌以及真菌中的根霉等。
、短乳杆菌(Lactobacillus brevis)、甘露醇乳杆菌(Lac. manitopoeum)、双歧杆菌以及真菌中的根霉等。.")
117
由于微生物代谢途径都不是单一的,所以代谢产物也并非像代谢途径描述的那么单纯,乳酸发酵不论同型发酵还是异型发酵,产物并没有不可逾越的界限。在微生物分类学研究中,通常把1分子葡萄糖发酵生成的乳酸少于1.8分子,同时产生较多的乙醇、二氧化碳、甘油、乙酸、甘露醇等产物的乳酸菌称为异型乳酸发酵菌。异型乳酸发酵已被广泛应用到生理活性饮料的生产工艺中。

118
三、脱氧酮糖酸途径(ED途径)与细菌酒精发酵
脱氧酮糖酸途径又称ED途径。微生物除通过糖酵解途径、三羧酸循环、戊糖磷酸途径等对葡萄糖进行降解外,某些微生物还能通过其他途径进行降解。有些专性厌氧菌只具有ED途径进行单糖的分解代谢,以维持生长。这些微生物能够利用途径中生成的3-磷酸甘油醛沿戊糖磷酸途径的可逆反应与6-磷酸果糖结合,生成5-磷酸核糖、4-磷酸赤藓糖及其它中间产物,进而为核苷酸、氨基酸的生物合成及核酸、蛋白质等生物大分子与辅酶的合成提供前体物质。

120
少数兼性厌氧细菌利用葡萄糖经脱氧酮糖酸途径进行酒精发酵(图10-8)。细菌酒精发酵过程中,1分子葡萄糖经ED途径的两个产能反应,共生成2分子ATP,补偿开始消耗的一个,净得1个ATP,总反应式为:
。细菌酒精发酵过程中,1分子葡萄糖经ED途径的两个产能反应,共生成2分子ATP,补偿开始消耗的一个,净得1个ATP,总反应式为:")
121
第五节 糖异生作用 糖异生作用是指从非糖前体(包括丙酮酸、乳酸、柠檬酸循环中间体、大部分氨基酸的碳骨架及甘油等)合成葡萄糖。由非糖物质转变为葡萄糖的途径是由丙酮酸开始的,经一系列反应通过草酰乙酸形成葡萄糖。糖异生作用的主要部位是肝脏,肾脏也有但相差甚远,有非常少的糖异生作用存在于脑或肌肉中。糖异生作用是在饥饿或急需葡萄糖的情况下才产生的,其功用是保证血糖糖度的相对稳定。
合成葡萄糖。由非糖物质转变为葡萄糖的途径是由丙酮酸开始的,经一系列反应通过草酰乙酸形成葡萄糖。糖异生作用的主要部位是肝脏,肾脏也有但相差甚远,有非常少的糖异生作用存在于脑或肌肉中。糖异生作用是在饥饿或急需葡萄糖的情况下才产生的,其功用是保证血糖糖度的相对稳定。")
122
一、糖异生途径

123
二、糖酵解与糖异生的相互调节 (1)磷酸果糖激酶(PFK)和1,6-二磷酸果糖酶的调节 在代谢过程中,当AMP的水平高时,表明需要合成更多的ATP。AMP激发PFK,加快糖酵解的速率,同时抑制1,6-二磷酸果糖酶的活性,关闭糖异生作用。反之,当ATP和柠檬酸水平高时,表明不需要制造更多的ATP,高水平的ATP和柠檬酸抑制PFK,降低糖酵解的速率,同时柠檬酸激发1,6-二磷酸果糖酶的活性,加快糖异生作用的速率。

124
2)丙酮酸激酶、丙酮酸羧化酶和磷酸烯醇式丙酮酸(PEP)羧激酶的调节
高水平的ATP和丙氨酸抑制丙酮酸激酶,因此当ATP和生物合成的中间产物充足时,糖酵解被抑制,同时高水平的乙酰-CoA活化丙酮酸羧化酶,有助于糖异生作用的进行;反之,当细胞的供能状态低迷时,高水平的ADP抑制丙酮酸羧化酶和PEP羧激酶的活性,关闭糖异生作用,糖酵解作用开始。1,6-二磷酸果糖激活丙酮酸激酶,当糖酵解加速时,丙酮酸激酶的活性相应提高。

126
思考题 1. 磷酸戊糖途径的生物学意义是什么? 2.一分子葡萄糖完全氧化可生成30个或32个ATP,写出有关的代谢反应。
1. 磷酸戊糖途径的生物学意义是什么? 2.一分子葡萄糖完全氧化可生成30个或32个ATP,写出有关的代谢反应。 3. 淀粉酶一般可分为几种?它们的作用特点如何? 4. 以EMP途径为基础的发酵有哪些?为什么不同微生物发酵葡萄糖产生的发酵产物不一样?用什么方法可以使酵母法酵葡萄糖积累甘油? 5. 何谓乙醛酸循环?生理意义如何?

127
6. 何谓丙酮酸羧化支路?生理意义如何? 7. 何谓单磷酸己糖途径?生理意义如何? 8. 淀粉发酵生产柠檬酸途径如何?简述实现柠檬酸大量积累的生化机制? 9. 若以14C标记葡萄糖的C3作为酵母的底物,经发酵产生CO2和乙醇,试问14C将在何处发现? 10. 在严格的厌氧条件下酒精发酵过程中,使用放射性标记的碳源进行示踪原子实验。如果葡萄糖的第一个碳用14C标记,那么14C将出现在产物乙醇的哪个位置上?在起始的葡萄糖分子的哪个位置上标记14C,才能使乙醇发酵释放出的CO2都是14C 标记的14CO2。

128
11. 电子传递链和氧化磷酸化之间有何关系? 12. 什么是磷/氧比(P/O比),测定磷/氧比有何意义? 13. 计算从丙酮酸合成葡萄糖需提供多少高能磷酸键? 14. 葡萄糖、戊糖磷酸途径和葡萄糖异生途径之间如何联系? 15. 为什么有人不能耐受乳糖?而乳婴却靠乳汁维持生命? 16. 酵母可以依赖葡萄糖厌氧或有氧生长,试解释当一直处于厌氧环境中的酵母细胞暴露于空气中时,葡萄糖的消耗速率为什么会下降?

129
17. 葡萄糖的合成过程同分解过程是否为可逆反应?
18. 一般说糖酵解是无氧分解,三羧酸循环是有氧分解,严格说,这种说法是否正确?为什么? 19. 糖代谢与发酵工业有何联系? 20. EMP、HMP、PK、ED途径的关键酶是什么?这些途径间彼此有什么联系? 21.试比较细菌酒精发酵与酵母酒精发酵的异同。













 反应部位 器官,细胞内定位 生理意义 代谢调节 主要调节点,主要变构抑制剂、变构激活剂 各代谢途径之间的联系和调控.>")



 合成代谢------需要能量 物质代谢 能量代谢>")














